
(3)抗有害物质的损伤作用:英膜处于菌细胞的最外层,有保护菌体避免和减少受 溶菌酶、补体、抗菌抗体、抗菌药物等有害物质的损伤作用。 鞭毛许多细菌,包括所有的弧菌和螺菌,约半数的杆菌和个别球菌,在菌体上附有 细长并呈波状弯曲的丝状物,少仅1一2根,多者达数百。这些丝状物称为鞭毛(gd km),是细南的运动器官。鞭毛长5~20um,直径12~30nm,需用电子显徽镜观察(图 1-11),或经特殊染色法使鞭毛增粗后才能在普通光学显微镜下看到(图1-12)。 困12伤寒沙门菌的穰毛 毛染色 ×1900) 根据鞭毛的数量和部位,可将鞭毛菌分成 四类(图1-13)。①单毛菌(monotrichate):只 有一根鞭毛,位于菌体一端,如徭乱弧菌; ②双毛菌(amphitrichate):菌体两端各有一根 鞭毛,如空肠弯曲菌;③丛毛菌 《ophotrichate):菌体一端或两端有一丛鞭毛, 如铜绿假单胞菌;④周毛曹(peritrichate):菌 丛毛道 体周身遍布许多鞭毛,如伤寒沙门菌。 双毛 1,鞭毛的结构鞭毛自细胞膜长出,游离 单毛幽 于菌细胞外,由基础小体、钩状体和丝状体三 图1-13细菌鞭毛的类型 个部分组成(图1-14)。 ()基础小体(basal bdy):位于鞭毛根都,帐在细胞壁和细胞膜中。革兰阴性题糠毛的基酷小 体由一根圆柱、两对同心环和输出装置组成。其中,一对是M(membrane)环和S(supramembrane 环,附着在细胞膜上;另一对是P(peptidoglyc)环和L(lipopolychride)环,附着在细胞壁的肽 聚箭和外膜的脂多糖上。基础小体的著底部是鞭毛的输出装置(exportppu),位于细购膜内面的 细胞质内。基底部圈柱体周阙的发动器(motor)为鞭毛运动提供能量,近旁的开关(switch)决定鞭 毛转动的方向。革兰阳性菌的细胞壁无外膜,其鞭毛只有M、S一对同心环。 -21-

外膜 肽震椅 R. @ 7@ 细膜 发动 图14大肠埃希菌毛根部结构模式图 (2)钩状体(0k):位于鞭毛伸出菌体之处,呈约90'的钩状弯曲。鞭毛由此转弯向外伸出,成 为丝状体。 (3)丝状体(lament):呈纤丝状,伸出于菌体外,是由鞭毛蛋白(gein)紧密排列并缠绕而 成的中空管状结构。丝状体的作用犹如船舶或飞机的旋桨推进器。鞭毛蛋白是一种弹力纤维蛋白 其氨基酸组成与骨格肌中的肌动蛋白相似,可能与鞭毛的运动有关。 豫毛是从尖端生长,在曹体内形成的镰毛蛋白分子不新地添加到擦毛的末端。若用机械方法去降 鞭毛,新的鞭毛很快合成。3一6min内恢复动力。各菌种的機毛蛋白结构不同,具有高度的抗原性, 称为鞭毛(H)抗原。 2.鞭毛的功能具有鞭毛的细菌在液体环境中能自由游动,速度迅速,如单鞭毛 的震乱弧菌每秒移动可达55m,周毛菌移动较慢,每秒25一30um。细菌的运动有化 学趋向性,常向营养物质处前进,而逃离有害物质。 有些细菌的鞭毛与致病性有关。例如雷乱弧菌、空肠弯曲菌等通过活泼的鞭毛运动穿透 小肠粘膜表面覆盖的粘液层,使菌体粘附于肠粘膜上皮细胞,产生毒性物质导致病变的发生。 根据鞭毛菌的动力和鞭毛的抗原性,可用以鉴定细菌和进行细菌分类 细菌是由鞭毛发动器将跨膜质子梯度中贮存的化学能转变为鞭毛转动所需的能量,周浆间陈中的 质子(什)通过鞭毛发动器流人细胞质内。有少数细药管用的离子梯度供给廉毛转动的能量。在 这个过程中,由跨膜质子梯度或钠离子梯度构成质子动力(proton motive force)。鞭毛发动器能够顺 时钟或逆时钟方向转动,从而决定细菌游动的方向。当发动器逆时钟方向转动时,鞭毛的丝状体结合 成一束拖在蓝体后,推动细菌向前进(:若发动器呈顺时钟方向转动,束状丝状体松开,细菌停 顿或向相反方向游动(tumble)。平时,细菌以这两种方式交替游动,称为随意移动(random walk。 细菌的运动具有方向性,受环境因素的影响极大。菌细胞上有众多的特异信号受体(sglr心 c印o),能接受不同的理化和生物学刺激而作出相应反应。例如大肠埃希菌细胞膜上的特异性糖结合 受体。既能赛觉化学趋化信号,也参与该物质的运输。如果遇到吸引性刺徽时、细萄就会暂时性抑制 发动器的顺时钟方向转动,使菌体向吸引物移动:反之,遇到有岩物质时,也会增强发动器的顺时钟 方向转动,于是细菌背离有害物运动以保存自己。 -22-
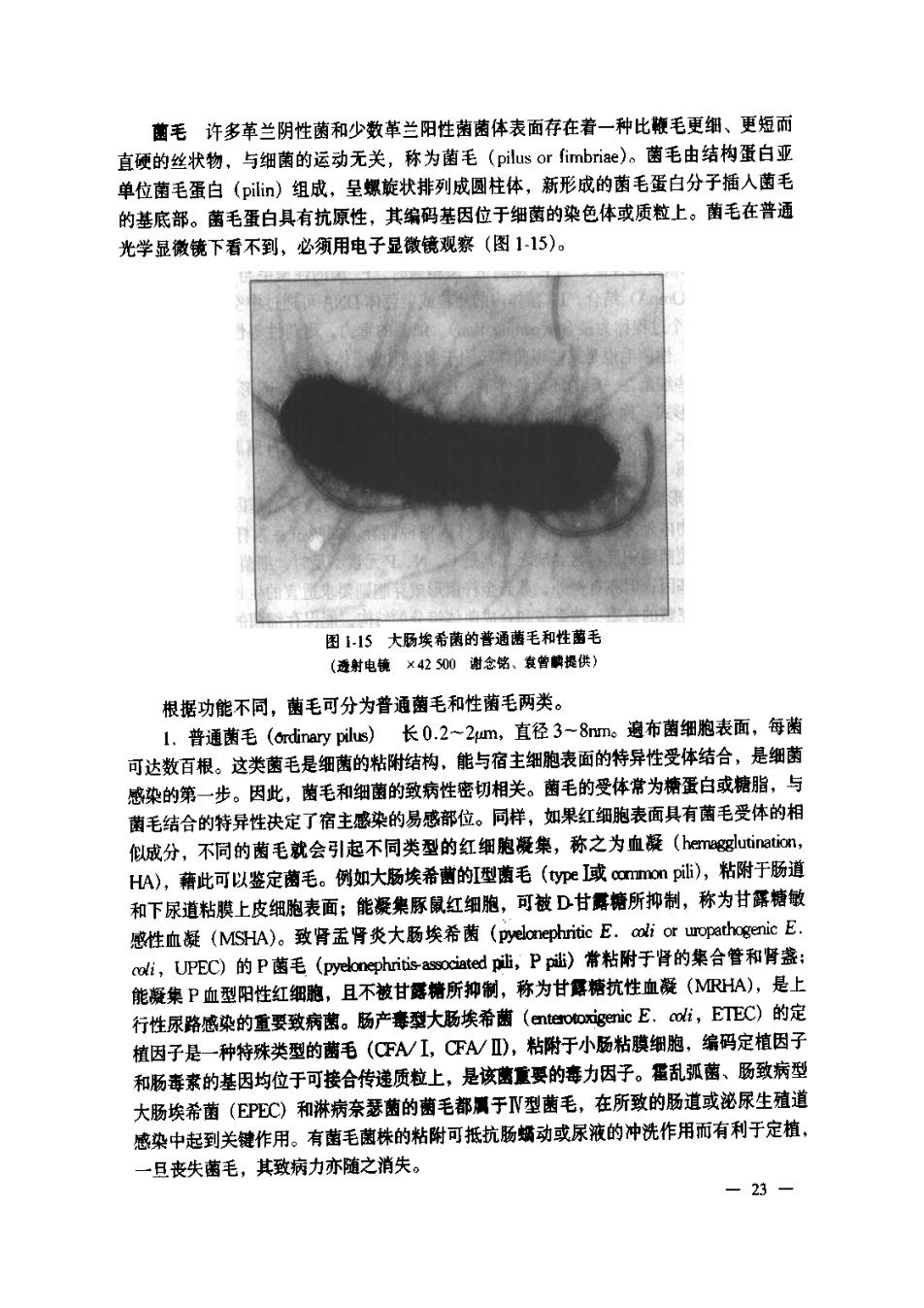
菌毛许多革兰阴性菌和少数革兰阳性菌菌体表面存在着一种比鞭毛更细、更短而 直硬的丝状物,与细菌的运动无关,称为菌毛(pilus or fimbrie)。菌毛由结构蛋白亚 单位菌毛蛋白(pil)组成,呈螺旋状排列成圆柱体,新形成的菌毛蛋白分子插入菌毛 的基底部。菌毛蛋白具有抗原性,其编码基因位于细菌的染色体或质粒上。菌毛在普通 光学显微镜下看不到,必须用电子显微镜观察(图115)。 困5大肠埃希菌的普通蓝毛和牲蕾毛 (透射电镜×42500谢念铭、熹曾膜提供 根据功能不同,菌毛可分为普通菌毛和性菌毛两类。 L.普通菌毛(rdiary pilus)长0.2~2m,直径3-8nm。遍布菌细胞表面,每菌 可达数百根。这类菌毛是细菌的粘附结构,能与宿主细胞表面的特异性受体结合,是细菌 感染的第一步。因此,曹毛和细菌的致病性密切相关。菌毛的受体常为糖蛋白或糖脂,与 菌毛结合的特异性决定了宿主感染的易感部位。同样,如果红细胞表面具有菌毛受体的相 似成分,不同的菌毛就会引起不同类型的红细胞凝巢,称之为血凝(hemagglutination, H),藉此可以鉴定菌毛。例如大肠埃希曹的I型菌毛(ye或common pi),粘附于肠道 和下尿道粘膜上皮细胞表面;能凝樂豚鼠红细胞,可被D甘舞糖所抑制,称为甘露糖敏 感性血凝(MSHA)。致肾盂肾炎大肠埃希(pyelnephritic E.cli or uropethogenic E. aoli,UPEC)的P蓝毛(pyeknepripil,P)常粘附于肾的集合管和肾盏: 能凝集P血型阳性红细胞,且不被甘露精所抑制,称为甘孱糖抗性血凝(MRHA),是上 行性尿路感染的重要致病曹。肠产喜型大肠埃希菌(nterigenic E.cot,ETEC)的定 植因子是一种特殊类型的菌毛(FA/I,CTA/),粘附于小肠粘膜细胞,编码定植因子 和肠毒素的基因均位于可接合传递质粒上,是该曹重要的毒力因子。乱弧菌、肠致病型 大肠埃希菌(EPEC)和淋病奈瑟菌的菌毛都于V型菌毛,在所致的肠道或泌尿生殖道 感染中起到关键作用。有菌毛菌株的粘附可抵抗肠娇动或尿液的冲洗作用而有利于定植, 一旦丧失菌毛,其致病力亦随之消失。 23

在革兰阳性球菌中,A群链球菌的菌毛与M蛋白和LTA结合在一起,介导该菌与 宿主粘模上皮细胞的粘附。 2.性毛(sxpilus) 仅见于少数革兰阴性菌。数量少,一个菌只有1一4根。 比普通菌毛长而粗,中空呈管状。性菌毛由一种称为致育因子(fertility factor,Ffac to)的质粒编码,故性菌毛又称F菌毛。带有性菌毛的细菌称为F*菌或雄性曹,无性 菌毛者称为F~菌或雄性菌。当F+菌与F~菌相遇时,F+菌的性菌毛与F菌相应的性 菌毛受体(如OmpA)结合,F+菌体内的质粒或染色体DNA可通过中空的性菌毛进人 F菌体内,这个过程称为接合(conjugation)。细菌的毒力、耐药性等性状可通过此方 式传递。此外,性菌毛也是某些噬菌体吸附于菌细胞的受体。 芽胞某些细菌在一定的环境条件下,能在菌体内部形成一个圆形或卵圆形小体, 是细菌的休眠形式,称为内芽胞(endospore),简称芽胞(spore),以别于真菌在菌体 外部形成的孢子。产生芽胸的细菌都是革兰阳性菌,重要的有芽胞杆菌属(炭疽芽胞杆 菌等)和梭菌属(破伤风梭菌等)。 1.芽胞的形成与发芽细菌形成芽胞的能力是由菌体内的芽胞基因决定的。芽胞 一般只是在动物体外才能形成,其形成条件因菌种而异。如炭疽芽胞杆菌在有氧下形 成,而破伤风梭菌则相反。营养缺乏尤其是C、N、P元豪不足时,细菌生长繁殖诚速、 启动芽胞形成基因;但亦有例外,苏云金杆菌形成芽胞则要求适宜的生长条件。 芽胞带有完整的核质、酶系统和合成菌体组分的结构,能保存细菌的全部生命必需 物质。芽胞形成后,菌体即成为空壳,有些芽 胞可从菌体脱落游离。 芽胞折光性强,壁厚,不易着色。染色时 需经媒染、加热等处理。芽胞的大小、形状、 位置等随菌种而异,有重要的鉴别价值(图1 16)。例如炭疽芽胞杆菌的芽胞为卵圆形、比 116细芽胞的形态、大小和位量 图117胶伤风梭菌芽胞 (透射电镜×21000谢念帖提供) 24

菌体小,位于菌体中央;破伤风梭菌芽胞呈圆形,比蓝体大,位于顶端,状如鼓槌(图 1-17):肉毒梭菌芽胞亦比菌体大,位于次极端。 芽胞形成在形态学上可分I一七个期,全程6一8 ,始于对数生长期末,菌细胞膜进行性地内陷 性生长,逐渐形成双层膜结构,包被核质成为芽胞的核心。细胞膜又能合成特殊物质,在内膜和外膜间 形成芽胞壁和皮质。在外膜外围再形成芽胞壳和芽胞外衣 成热的芽胞具有多层膜结构(困118)。芽胞核心 (o©r©)是芽胞的原生质体,含有细暂原有的核质和核糖体、 隋类等主要生命基质。核心的外层依次为内膜、芽胞壁、皮 芽胞外 质,外限、芽胞壳和芽胞外衣,将其层层包裹,成为坚实的 球体。内膜和外膜由原来的细胞膜形成。芽胞壁(sp wl)含肽聚糖,发芽后成为细菌的细胞壁。皮质(cnex 是芽胞包膜中最厚的一层,由一种特殊的肽聚精组成。芽跑 壳(a)是一种类似角蛋白的疏水性蛋白质,致密无通透 性,能抗化学药物进人,并增强对紫外线照射的抵抗力。有 些细菌芽胞还有一层兢松的芽胞外衣(exceporium),含有脂 图1-18细菌芽胞的结构 蛋白和糖类。 芽胞形成后,若由于机械力、热、pH改变等刺激作用,破坏其芽胞壳,并供给水 分和营养,芽胞可发芽,形成新的菌体。 一个细菌只形成一个芽胞,一个芽胞发芽也只生成一个茵体,细菌数量并未增加, 因而芽胞不是细菌的繁殖方式。与芽胞相比,未形成芽胞而具有繁殖能力的菌体可称为 靠殖体(vegetative form). 细菌的芽胞发芽(en)成紫殖体的过程,可分为活化(activation】、启动(initi)和 长出(outgrowth)三个连续阶段。整个过程大约需要90min。热刺漱(如60C1h或85℃5mim)和 p州降低均可活化芽胞发芽、L丙氨酸、葡萄糖、肌苷和腺苷均为启动剂。芽胞壳经活化后,其富含 二硫健的蛋白构型变化,引起渗透性改变,致使阳离子渗人,细胞膜脂质活性增强,并启动电子传递 硅。同时。随着水分渗人。芽胞特有成分吡院二骏酸钙、皮质肽豪糖和芽胞壳物质等大量降解,使芽 胞通透性加强,耐热、抗射等特性消失。由于代谢活性和呼吸增强,生物合成加速,顺序为RNA 蛋白质、脂质,最后是DA。继而芽胞核心体积增大、皮质膨松、芽胞壳破裂,芽管长出并逐渐长 大、发育成新的繁殖体细胞。 2.芽胞的功能细随的芽胞对热力、干燥、辐射、化学消毒剂等理化因素均有强 大的抵抗力。一般细菌繁殖体在80℃水中迅速死亡,而有的细菌芽胞可耐100℃沸水数 小时。被炭疽芽胞杆菌芽胞污染的草原,传染性可保持20一30年。 细菌芽胞并不直接引起疾病,仅当发芽成为繁殖体后,才能迅速大量繁殖而致病 例如土壤中常有破伤风梭菌的芽胞,一旦外伤深部创口被泥土污染,进入伤口的芽胞在 适宜条件下即可发芽成繁殖体再产毒致病。 被芽胞污染的用具、敷料、手术器械等,用一般方法不易将其杀死,杀灭芽胞最可 靠的方法是高压蒸气灭菌。当进行消毒灭菌时,应以芽胞是否被杀死作为判断灭菌效果 的指标。 细菌芽胞抵抗力强的原因,可能与下列因素有关:①芽胞含水量少,约占繁殖体的40%,蛋白 -25-