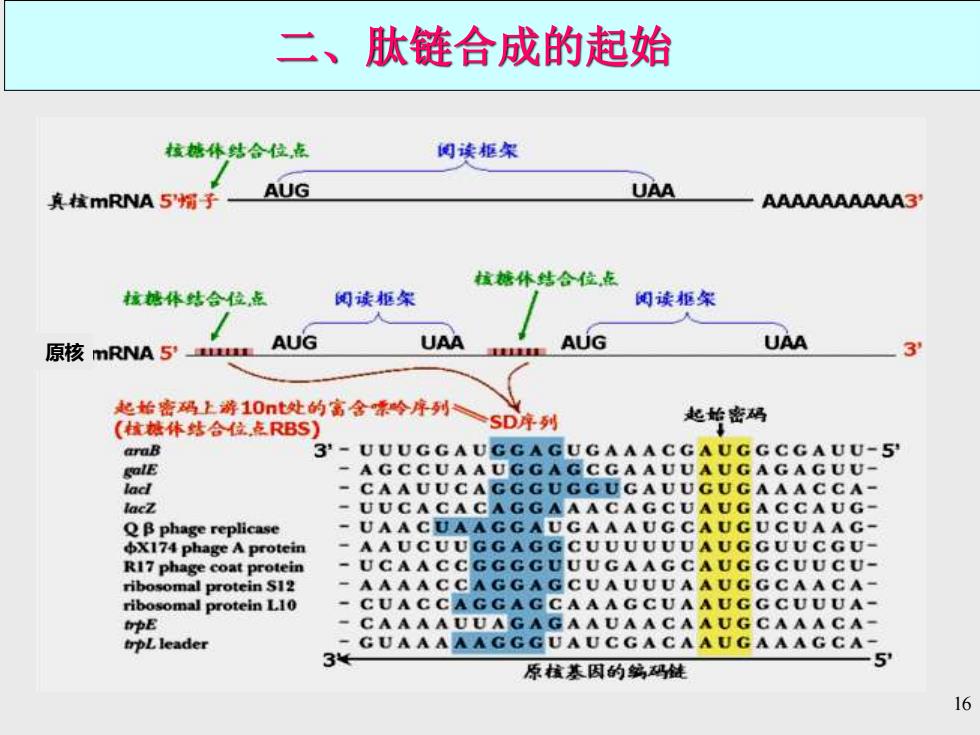
二、肽链合成的起始 核塘体结合位点 阅读拒宋 真核mRNA5帽子 AUG UAA AAAAAAAAAA3 核塘休结合位点 核糖体结合位,点 阅读柜架 阅读拒架 原核mRNA 5' AUG UAA 重直 AUG UAA 3 起指密码上游10nt处的富金价序列SD年列 起始密码 (植抽体结合位,点RBS araB 3-UUUGGAUGGAGUGAAACGAUGGCGAUU-5 golE -AGCCUAAUGGAGCGAAUUAUGAGAGUU- lacl -CAAUUCAGGGUGGUGAUUGUGAAACCA- lacZ -UUCACACAGGAAACAGCUAUGACCAUG- QB phage replicase -UAACUAAGGAUGAAAUGCAUGUCUAAG- X174 phage A protein -AAUCUUGGAGGCUUUUUUAUGGUUCGU- R17 phage coat protein -UCAACCGGGGUUUGAAGCAUGGCUUCU- ribosomal protein S12 -AAAACCAGGAGCUAUUUAAUGGCAACA- ribosomal protein L10 -CUACCAGGAGCAAAGCUAAUGGCUUUA- tpE -CAAAAUUAGAGAAUAACAAUGCAAACA- trpLleader 3GUAAAAAGGGUAUCCACAAUCAAACCA- 原罐基因的鹤码链 5 16
16 原核 二、肽链合成的起始

16 S rRNA与SD序列的配对 E.colitrpA (5)AG C A CG A GGG GA AA U C U G A U GG AA C G C U A C(3') E.coli ara B UUU GG AUGG A GU G AA A C GAUGG CG A U U G CA E.coli lac I C AA UU C A G GG U GG UG AA U G U O AAA CC A G U A 6X174 phage A protein AAUCUUGGAGGCUUUUUUAUGG UUCGUUCU 入phage cro A U G U A C U AA GG AG G UU G U A U GG AA C AA C G C Shine-Dalgarno序列; 起始密码子: 能与16SRNA时 能与fMet-RNAfMeti程对 (a) Shine.及Dalgarno等证明几乎所有原核生物 mRNA上都有一个S'-AGGAGGU3'序列,这 16SrRNA 16 SrRNA对 个富嘌呤区与30S亚基上16 SrRNA3'末端的 的3端 原核mRNA 富嘧啶区5-GAUCACCUCCUUA3相互补。 的SD序列的 (5)GA UUC C UAG GAG GUU U G A CC UAUGC GAG CUUU UAGU(3 识别和结合 (b) ◆原核生物的mRNA存在富含梁吟碱基的SD序列 ◆核糖体小亚基上的16 SrRNA3'端的富含蜜啶序列可与SD序列配对结合 ◆真核生物不存在SD序列,但存在5?帽子和3'PoyA尾巴结构 ◆真核生物蛋白质合成的启动需要帽于结合蛋白
17 Shine及Dalgarno等证明几乎所有原核生物 mRNA上都有一个5’-AGGAGGU-3’序列,这 个富嘌呤区与30S亚基上16S rRNA 3’末端的 富嘧啶区5’-GAUCACCUCCUUA-3’相互补
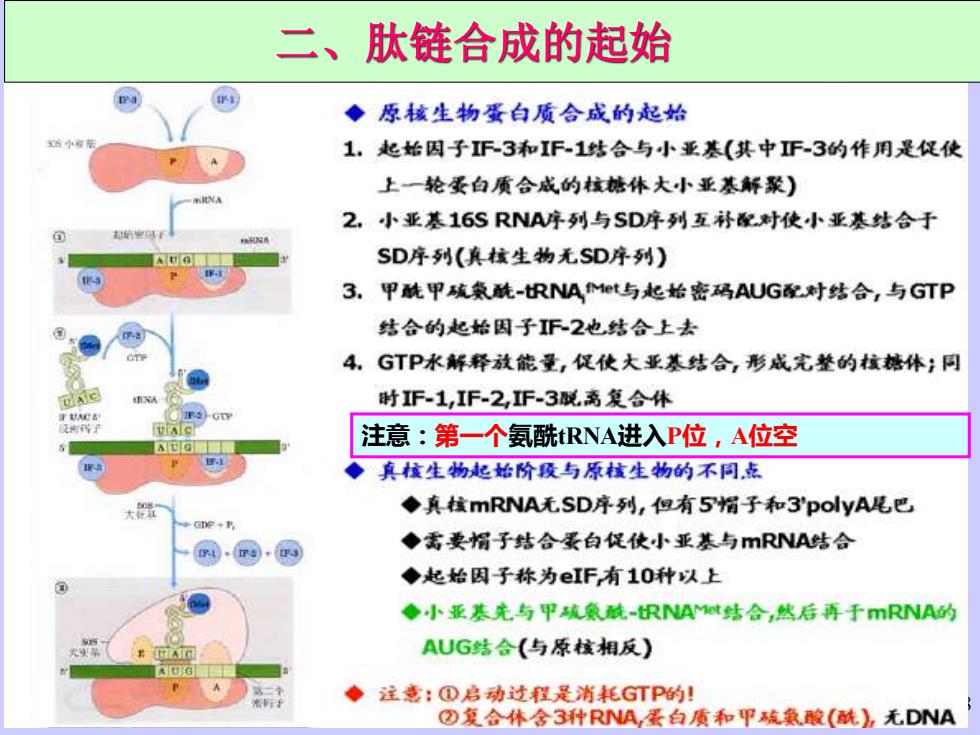
二、肽链合成的起始 ◆原核生物蛋白质合成的起始 1,起始因子F-3和IF-1结合与小亚基(其中F-3的作用是促使 上一轮蛋白质合成的核糖休大小亚基解聚) A 2,小亚基16SRNA序列与SD序列互补配对使小亚基结合于 SD序列(真核生物无SD序列) 3.甲酰甲硫数酰-RNAt与起始密码AUG对结合,与GTP 结合的起始因子F2也,结合上去 4 GTP水解释放能量,促使大亚基结合,形成完整的核糖体;同 时1F-1,IF-2,F-3脱离复合休 注意:第一个氨酰tRNA进入P位,A位空 ◆真核生物起始阶段与原核生物的不同志 大 ◆真核mRNA无SD序列,但有5帽子和3'polyA昆巴 e+ 问·同 ◆需要帽子结合景白促使小亚基与mRNA结合 ◆起始因子称为eIF有10种以上 ◆小亚基先与甲硫象酰-RNA结合,然后再于mRNA的 AUG结合(与原核相反) ◆注意:①启动过程是消耗GTP的! ②复合体含3种RNA,蛋白质和甲硫魚酸(酰),无DNA
18 二、肽链合成的起始 注意:第一个氨酰tRNA进入P位,A位空

真核和原核细胞参与翻译的蛋白质因子 阶段 原核 真核 功 能 IF1 IF2 eIF2 参与起始复合物的形成 IF3 eIF3、elF4C 起始 CBPI 与mRNA帽子结合 eIF4A B F 参与寻找第一个AUG eIF5 协助eF2、eF3、eF4C的释放 elF6 协助60S亚基从无活性的核糖体上解离 EF-Tu eEFla 协助氨酰-tRNA进入核糖体 延长 EF-Ts eEF1βy 帮助EF-Tu、eEF1o周转 EF-G eEF2 移位因子 RF-1 终止 eRF 释放完整的肽链 RF-2
19 阶段 原核 真核 功 能 IF1 IF2 eIF2 参与起始复合物的形成 IF3 eIF3、eIF4C 起始 CBP I 与mRNA帽子结合 eIF4A B F 参与寻找第一个AUG eIF5 协助eIF2 、 eIF3、eIF4C的释放 eIF6 协助60S亚基从无活性的核糖体上解离 EF-Tu eEF1 协助氨酰-tRNA进入核糖体 延长 EF-Ts eEF1 帮助EF-Tu 、 eEF1周转 EF-G eEF2 移位因子 RF-1 终止 eRF 释放完整的肽链 RF-2 真核和原核细胞参与翻译的蛋白质因子

。 细菌核糖体上一般存在三个与氨基酰 tRNA结合的位点:即A位点(aminoacyl site),P位点(peptidyl site)和E位点 (Exit site)。 ·只有Met-tRNAfMet能与第一个P位点相结 合,其它所有RNA都必须通过A位点到达 P位点,再由E位点离开核糖体。 20
20 • 细菌核糖体上一般存在三个与氨基酰- tRNA结合的位点: 即A位点(aminoacyl site),P位点(peptidyl site)和E位点 (Exit site)。 • 只有fMet-tRNAfMet能与第一个P位点相结 合,其它所有tRNA都必须通过A位点到达 P位点,再由E位点离开核糖体