
第一节概述 脂类是机体内的一类有机大分子物质,它包括范围很广,其化学结构有很大差异,生理功能各不相同,其共同理化性质是不溶于水而溶于 有机溶剂。 一、脂类的分类及其功能 脂类分为两大类,即脂肪(fat)和类脂(lipids) (一)脂肪:即甘油三脂或称之为脂酰甘油(triacylglycerol),它是由1分子甘油与3个分子脂肪酸通过酯键相结合而成。人体内脂肪酸种类很 多,生成甘油三脂时可有不同的排列组合,因此,甘油三脂具有多种形式。贮存能量和供给能量是脂肪最重要的生理功能。1克脂肪在体内完 全氧化时可释放出38kJ(9.3kca),比1克糖原或蛋白质所放出的能量多两倍以上。脂防组织是体内专门用于贮存脂肪的组织,当机体需要时, 脂肪组织中贮存在脂肪可动员出来分解供给机体能量。此外,脂肪组织还可起到保持体温,保护内脏器官的作用。 (二)类脂:包括磷脂(phospholipids),糖脂(glycolipid)和胆固醇及其酯(cholesterol and cholesterol ester))三大类。磷脂是含有磷酸的脂 类,包括由甘油构成的甘油磷脂(phosphoglycerides)和由鞘氨醇构成的鞘磷脂(sphingomyelin)。糖脂是含有糖基的脂类。这三大类类脂是生物 膜的主要组成成分,构成疏水性的“屏障”(barrier),分隔细胞水溶性成分和细胞器,维持细胞正常结构与功能。此外,胆固醇还是脂肪酸盐和维 生素D3以及类固醇激素合成的原料,对于调节机体脂类物质的吸收,尤其是脂溶性维生素(A,D,E,K)的吸收以及钙磷代谢等均起着重要作 用。 二、脂类的消化和吸收 正常人一般每日每人从食物中消化60?50克的脂类,其中甘油三脂占到90%以上,除此以外还有少量的磷脂、胆固醇及其酯和一些游离脂 防酸(free fattyacids)。食物中的脂类在成人口腔和胃中不能被消化,这是由于口腔中没有消化脂类的酶,胃中虽有少量脂防酶,但此酶只有在 中性PH值时才有活性,因此在正常胃液中此酶几乎没有活性(但是婴儿时期,胃酸浓度低,胃中PH值接近中性,脂肪尤其是乳脂可被部分消 化)。脂类的消化及吸收主要在小肠中进行,首先在小肠上段,通过小肠蠕动,由胆汁中的胆汁酸盐使食物脂类乳化,使不溶于水的脂类分散 成水包油的小胶体颗粒,提高溶解度增加了酶与脂类的接触面积,有利于脂类的消化及吸收。在形成的水油界面上,分泌入小肠的胰液中包含 的酶类,开始对食物中的脂类进行消化,这些酶包括胰脂肪酶(pancreatic lipase),辅脂酶(colipase),胆固醇酯酶(pancreatic cholesteryl ester hydrolase or cholesterol esterase)和磷脂酶A2(phospholipase A2). 食物中的脂肪乳化后,被胰脂肪酶催化,水解甘油三酯的1和3位上的脂肪酸,生成2·甘油一酯和脂肪酸。此反应需要辅脂酶协助,将脂 防酶吸附在水界面上,有利于胰脂酶发挥作用。 甘油三酯一一率防鹿2甘油一脂十2X脂防酸 食物中的磷脂被磷脂酶A2催化,在第2位上水解生成溶血磷脂和脂肪酸,胰腺分泌的是磷脂酶A2原,是一种无活性的酶原形成,在肠道被 胰蛋白酶水解释放一个6肽后成为有活性的磷脂酶A2催化上述反应。 磷脂一 网期德人溶血陕脂十脂脑酸 食物中的胆固醇酯被胆固醇酯酶水解,生成胆固醇及脂防酸。 胆固尊蜜一靡鹿胆固醇十脂肪酸 食物中的脂类经上述胰液中酶类消化后,生成甘油一酯、脂肪酸、胆固醇及溶血磷脂等,这些产物极性明显增强,与胆汁乳化成混合微团 (mixed micelles),这种微团体积很小(直径20nm),极性较强,可被肠粘膜细胞吸收. 脂类的吸收主要在十二指肠下段和盲肠。甘油及中短链脂肪酸(<=10C)无需混合微团协助,直接吸收入小肠粘膜细胞后,进而通过门静脉 进入血液。长链脂肪酸及其它脂类消化产∥/物随微团吸收入小肠粘膜细胞。长链脂肪酸在脂酰CoA合成酶(fattyacyl CoA synthetase)催化下, 生成脂酰CoA,此反应消耗ATP。 脂肪酸+HSCoA+ATP +首酰CoA+AMP 脂酰CoA可在转酰基酶(acyltransferase)作用下,将甘油一酯、溶血磷脂和胆固醇酯化生成相应的甘油三酯、磷脂和胆固醇酯。体内具有多 种转酰基酶,它们识别不同长度的脂肪酸催化特定酯化反应。 甘油一酯院CoN HSCoA甘油二酯晓CoA HSCoA甘泣三酯 溶血瞬脂n院CoA HSCA磷出 胆固醇脂酰CoA HSCoA 胆固醇酪
第一节 概 述 脂类是机体内的一类有机大分子物质,它包括范围很广,其化学结构有很大差异,生理功能各不相同,其共同理化性质是不溶于水而溶于 有机溶剂。 一、脂类的分类及其功能 脂类分为两大类,即脂肪(fat)和类脂(lipids) (一)脂肪:即甘油三脂或称之为脂酰甘油(triacylglycerol),它是由1分子甘油与3个分子脂肪酸通过酯键相结合而成。人体内脂肪酸种类很 多,生成甘油三脂时可有不同的排列组合,因此,甘油三脂具有多种形式。贮存能量和供给能量是脂肪最重要的生理功能。1克脂肪在体内完 全氧化时可释放出38kJ(9.3kcal),比1克糖原或蛋白质所放出的能量多两倍以上。脂肪组织是体内专门用于贮存脂肪的组织,当机体需要时, 脂肪组织中贮存在脂肪可动员出来分解供给机体能量。此外,脂肪组织还可起到保持体温,保护内脏器官的作用。 (二)类脂:包括磷脂(phospholipids),糖脂(glycolipid)和胆固醇及其酯(cholesterol and cholesterol ester)三大类。磷脂是含有磷酸的脂 类,包括由甘油构成的甘油磷脂(phosphoglycerides)和由鞘氨醇构成的鞘磷脂(sphingomyelin)。糖脂是含有糖基的脂类。这三大类类脂是生物 膜的主要组成成分,构成疏水性的“屏障”(barrier),分隔细胞水溶性成分和细胞器,维持细胞正常结构与功能。此外,胆固醇还是脂肪酸盐和维 生素D3以及类固醇激素合成的原料,对于调节机体脂类物质的吸收,尤其是脂溶性维生素(A,D,E,K)的吸收以及钙磷代谢等均起着重要作 用。 二、脂类的消化和吸收 正常人一般每日每人从食物中消化60?50克的脂类,其中甘油三脂占到90%以上,除此以外还有少量的磷脂、胆固醇及其酯和一些游离脂 肪酸(free fattyacids)。食物中的脂类在成人口腔和胃中不能被消化,这是由于口腔中没有消化脂类的酶,胃中虽有少量脂肪酶,但此酶只有在 中性PH值时才有活性,因此在正常胃液中此酶几乎没有活性(但是婴儿时期,胃酸浓度低,胃中PH值接近中性,脂肪尤其是乳脂可被部分消 化)。脂类的消化及吸收主要在小肠中进行,首先在小肠上段,通过小肠蠕动,由胆汁中的胆汁酸盐使食物脂类乳化,使不溶于水的脂类分散 成水包油的小胶体颗粒,提高溶解度增加了酶与脂类的接触面积,有利于脂类的消化及吸收。在形成的水油界面上,分泌入小肠的胰液中包含 的酶类,开始对食物中的脂类进行消化,这些酶包括胰脂肪酶(pancreatic lipase),辅脂酶(colipase),胆固醇酯酶(pancreatic cholesteryl ester hydrolase or cholesterol esterase)和磷脂酶A2(phospholipase A2)。 食物中的脂肪乳化后,被胰脂肪酶催化,水解甘油三酯的1和3位上的脂肪酸,生成2-甘油一酯和脂肪酸。此反应需要辅脂酶协助,将脂 肪酶吸附在水界面上,有利于胰脂酶发挥作用。 食物中的磷脂被磷脂酶A2催化,在第2位上水解生成溶血磷脂和脂肪酸,胰腺分泌的是磷脂酶A2原,是一种无活性的酶原形成,在肠道被 胰蛋白酶水解释放一个6肽后成为有活性的磷脂酶A2催化上述反应。 食物中的胆固醇酯被胆固醇酯酶水解,生成胆固醇及脂肪酸。 食物中的脂类经上述胰液中酶类消化后,生成甘油一酯、脂肪酸、胆固醇及溶血磷脂等,这些产物极性明显增强,与胆汁乳化成混合微团 (mixed micelles)。这种微团体积很小(直径20nm),极性较强,可被肠粘膜细胞吸收。 脂类的吸收主要在十二指肠下段和盲肠。甘油及中短链脂肪酸(<=10C)无需混合微团协助,直接吸收入小肠粘膜细胞后,进而通过门静脉 进入血液。长链脂肪酸及其它脂类消化产///物随微团吸收入小肠粘膜细胞。长链脂肪酸在脂酰CoA合成酶(fattyacyl CoA synthetase)催化下, 生成脂酰CoA,此反应消耗ATP。 脂酰CoA可在转酰基酶(acyltransferase)作用下,将甘油一酯、溶血磷脂和胆固醇酯化生成相应的甘油三酯、磷脂和胆固醇酯。体内具有多 种转酰基酶,它们识别不同长度的脂肪酸催化特定酯化反应
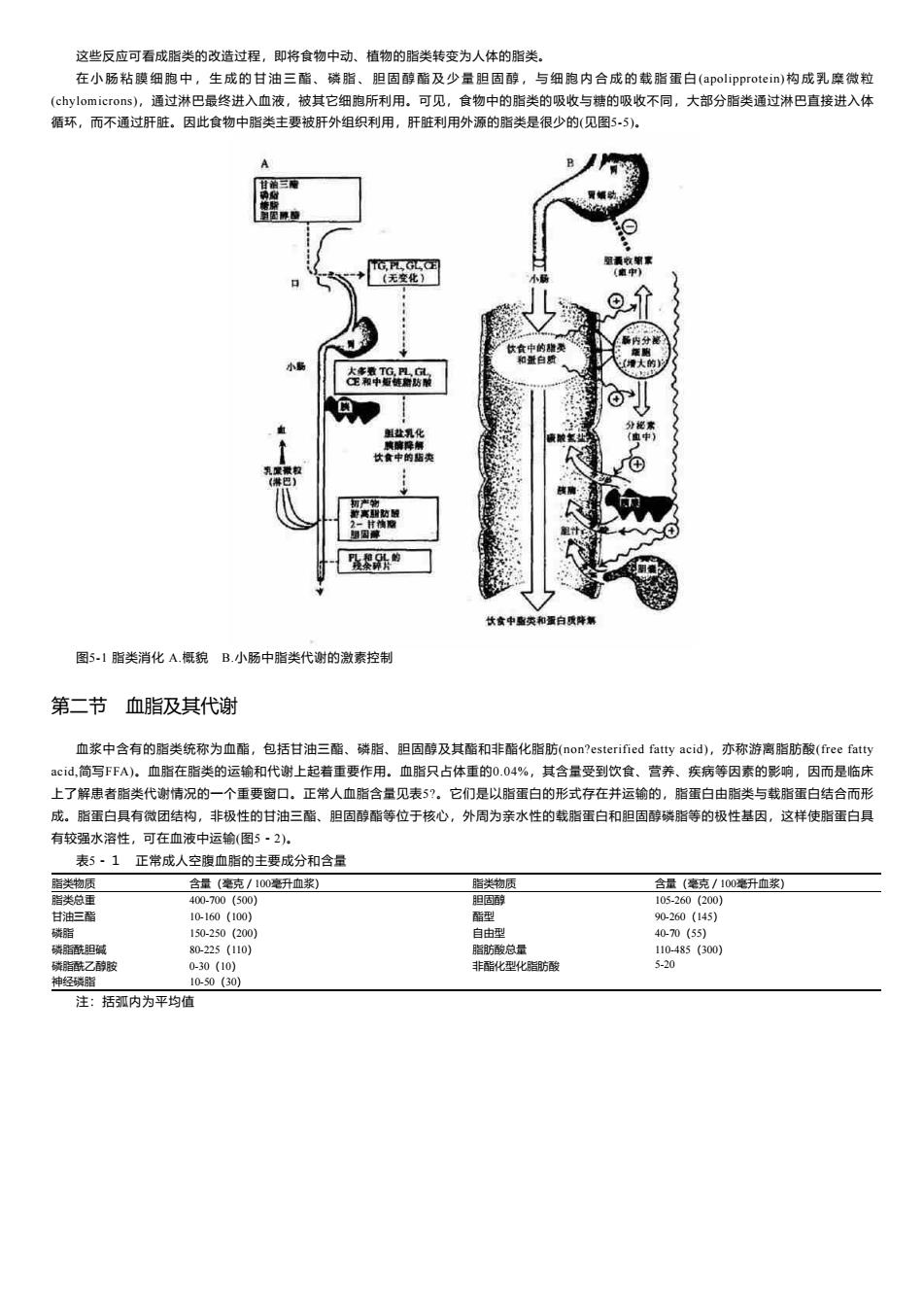
这些反应可看成脂类的改造过程,即将食物中动、植物的脂类转变为人体的脂类。 在小肠粘膜细胞中,生成的甘油三酯、磷脂、胆固醇酯及少量胆固醇,与细胞内合成的载脂蛋白(apolipprotein)构成乳糜微粒 (chylomicrons),通过淋巴最终进入血液,被其它细胞所利用。可见,食物中的脂类的吸收与糖的吸收不同,大部分脂类通过淋巴直接进入体 循环,而不通过肝脏。因此食物中脂类主要被肝外组织利用,肝脏利用外源的脂类是很少的(见图5-5)。 A (无变化》 休食中的维类 和置白 品 鲜让乳化 食中的见 离防酸 食中酯类和蛋白解 图5-1脂类消化A.概貌B.小肠中脂类代谢的激素控制 第二节 血脂及其代谢 血浆中含有的脂类统称为血酯,包括甘油三酯、磷脂、胆固醇及其酯和非酯化脂肪(non?esterified fatty acid),亦称游离脂肪酸(free fatty cid,简写FFA)。血脂在脂类的运输和代谢上起着重要作用。血脂只占体重的0.04%,其含量受到饮食、营养、疾病等因素的影响,因而是临床 上了解患者脂类代谢情况的一个重要窗口。正常人血脂含量见表5?。它们是以脂蛋白的形式存在并运输的,脂蛋白由脂类与载脂蛋白结合而形 成。脂蛋白具有微团结构,非极性的甘油三酯、胆固醇酯等位于核心,外周为亲水性的载脂蛋白和胆固醇磷脂等的极性基因,这样使脂蛋白具 有较强水溶性,可在血液中运输(图5·2). 表5·1正常成人空腹血脂的主要成分和含量 脂类物质 含量(毫克/100毫升血浆) 脂类物质 含量(毫克/100毫升血浆) 脂类总重 400-700(500) 胆固醇 105-260(200) 甘油三酯 10-160(100) 酯型 90-260(145) 磷脂 150-250(200) 自由型 40-70(55) 磷脂酰胆碱 80-225(110) 脂肪酸总量 110-485(300)】 磷脂酰乙醇胺 0-30(10) 非酯化型化脂肪酸 5-20 神经碳脂 10-50(30) 注:括弧内为平均值
这些反应可看成脂类的改造过程,即将食物中动、植物的脂类转变为人体的脂类。 在 小 肠 粘 膜 细 胞 中 , 生 成 的 甘 油 三 酯 、 磷 脂 、 胆 固 醇 酯 及 少 量 胆 固 醇 , 与 细 胞 内 合 成 的 载 脂 蛋 白 (apolipprotein) 构 成 乳 糜 微 粒 (chylomicrons),通过淋巴最终进入血液,被其它细胞所利用。可见,食物中的脂类的吸收与糖的吸收不同,大部分脂类通过淋巴直接进入体 循环,而不通过肝脏。因此食物中脂类主要被肝外组织利用,肝脏利用外源的脂类是很少的(见图5-5)。 图5-1 脂类消化 A.概貌 B.小肠中脂类代谢的激素控制 第二节 血脂及其代谢 血浆中含有的脂类统称为血酯,包括甘油三酯、磷脂、胆固醇及其酯和非酯化脂肪(non?esterified fatty acid),亦称游离脂肪酸(free fatty acid,简写FFA)。血脂在脂类的运输和代谢上起着重要作用。血脂只占体重的0.04%,其含量受到饮食、营养、疾病等因素的影响,因而是临床 上了解患者脂类代谢情况的一个重要窗口。正常人血脂含量见表5?。它们是以脂蛋白的形式存在并运输的,脂蛋白由脂类与载脂蛋白结合而形 成。脂蛋白具有微团结构,非极性的甘油三酯、胆固醇酯等位于核心,外周为亲水性的载脂蛋白和胆固醇磷脂等的极性基因,这样使脂蛋白具 有较强水溶性,可在血液中运输(图5-2)。 表5-1 正常成人空腹血脂的主要成分和含量 脂类物质 含量(毫克/100毫升血浆) 脂类物质 含量(毫克/100毫升血浆) 脂类总重 400-700(500) 胆固醇 105-260(200) 甘油三酯 10-160(100) 酯型 90-260(145) 磷脂 150-250(200) 自由型 40-70(55) 磷脂酰胆碱 80-225(110) 脂肪酸总量 110-485(300) 磷脂酰乙醇胺 0-30(10) 非酯化型化脂肪酸 5-20 神经磷脂 10-50(30) 注:括弧内为平均值
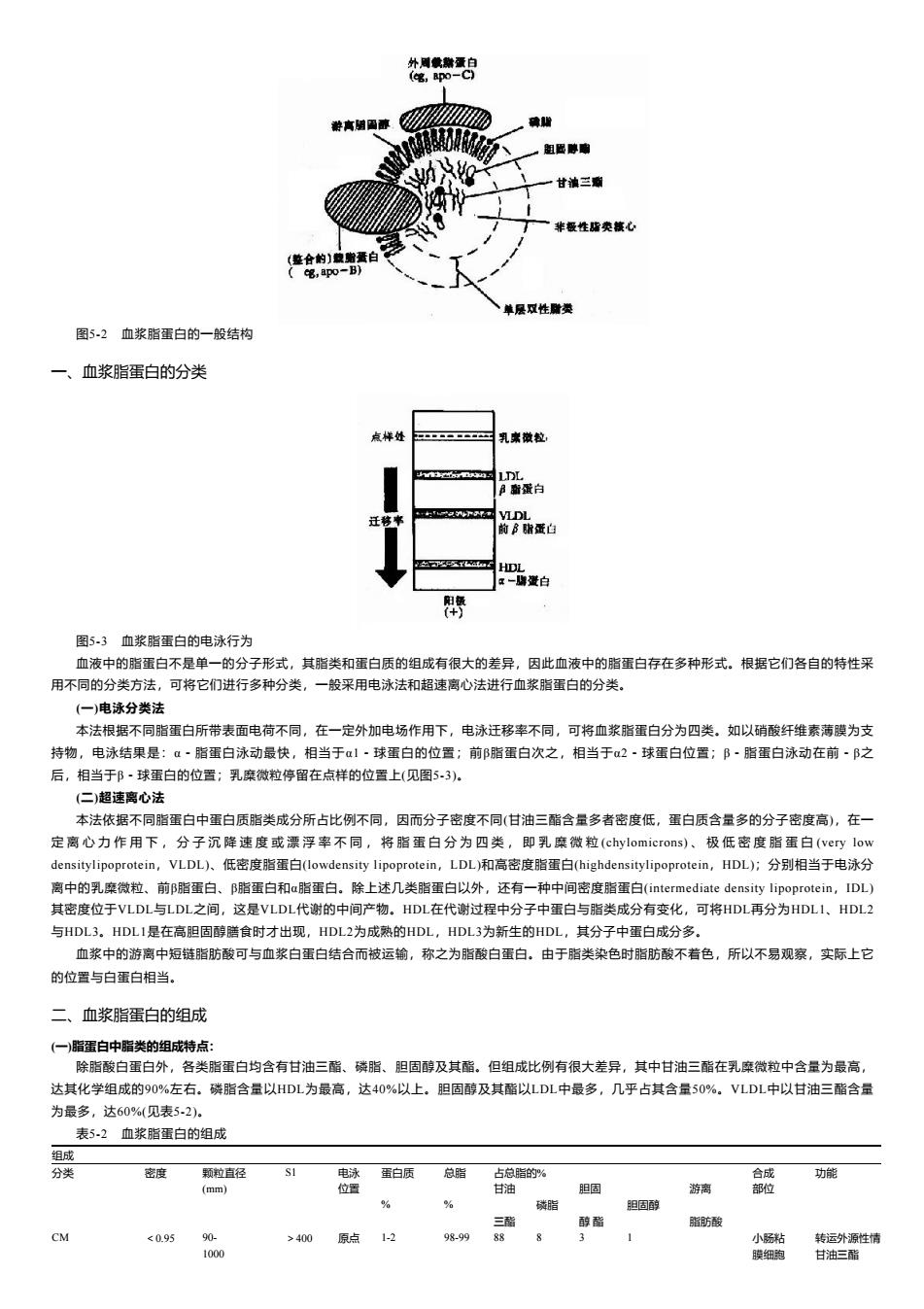
外周戴然蛋白 笔,pO-C 游高硝图藤 确 粗医脚扇 甘滇三聊 非极性隆类核心 (《整合的)煎赠蛋白 C吧,po-B) 单层双性附类 图5-2血浆脂蛋白的一般结构 一、血浆脂蛋白的分类 点样处 乳察做粒, LDL B雷蛋白 迁移率 VLDL 前B脂蛋白 HDL x-博爱白 帶 图5.3血浆脂蛋白的电泳行为 血液中的脂蛋白不是单一的分子形式,其脂类和蛋白质的组成有很大的差异,因此血液中的脂蛋白存在多种形式。根据它们各自的特性采 用不同的分类方法,可将它们进行多种分类,一般采用电泳法和超速离心法进行血浆脂蛋白的分类。 (一)电泳分类法 本法根据不同脂蛋白所带表面电荷不同,在一定外加电场作用下,电泳迁移率不同,可将血浆脂蛋白分为四类。如以硝酸纤维素薄膜为支 持物,电泳结果是:α·脂蛋白泳动最快,相当于α1·球蛋白的位置;前β脂蛋白次之,相当于α2·球蛋白位置;阝·脂蛋白泳动在前·B之 后,相当于阝·球蛋白的位置;乳糜微粒停留在点样的位置上(见图53)。 (二)超速离心法 本法依据不同脂蛋白中蛋白质脂类成分所占比例不同,因而分子密度不同(甘油三酯含量多者密度低,蛋白质含量多的分子密度高),在一 定离心力作用下,分子沉降速度或漂浮率不同,将脂蛋白分为四类,即乳糜微粒(chylomicrons)、极低密度脂蛋白(very low densitylipoprotein,VLDL)、低密度脂蛋白(lowdensity lipoprotein,LDL)和高密度脂蛋白(highdensitylipoprotein,HDL);分别相当于电泳分 离中的乳糜微粒、前B脂蛋白、B脂蛋白和a脂蛋白。除上述几类脂蛋白以外,还有一种中间密度脂蛋白(intermediate density lipoprotein,IDL) 其密度位于VLDL与LDL之间,这是VLDL代谢的中间产物。HDL在代谢过程中分子中蛋白与脂类成分有变化,可将HDL再分为HDL1、HDL2 与HDL3。HDL1是在高胆固醇膳食时才出现,HDL2为成熟的HDL,HDL3为新生的HDL,其分子中蛋白成分多。 血浆中的游离中短链脂肪酸可与血浆白蛋白结合而被运输,称之为脂酸白蛋白。由于脂类染色时脂防酸不着色,所以不易观察,实际上它 的位置与白蛋白相当。 二、血浆脂蛋白的组成 (一)脂蛋白中脂类的组成特点: 除脂酸白蛋白外,各类脂蛋白均含有甘油三酯、磷脂、胆固醇及其酯。但组成比例有很大差异,其中甘油三酯在乳糜微粒中含量为最高, 达其化学组成的90%左右。磷脂含量以HDL为最高,达40%以上。胆固醇及其酯以LDL中最多,几乎占其含量50%。VLDL中以甘油三酯含量 为最多,达60%(见表5-2)。 表5-2血浆脂蛋白的组成 组成 分类 密度 颗粒直径 电泳蛋白质 总脂 占总脂的% 合成 功能 (mm) 位置 甘油 胆圈 游离 部位 % % 磷脂 胆因醇 三酯 醇酯 脂肪酸 CM <0.95 90- >400 原点1-2 98-99 88 8 小肠粘 转运外源性情 1000 膜细跑 甘油三酯
图5-2 血浆脂蛋白的一般结构 一、血浆脂蛋白的分类 图5-3 血浆脂蛋白的电泳行为 血液中的脂蛋白不是单一的分子形式,其脂类和蛋白质的组成有很大的差异,因此血液中的脂蛋白存在多种形式。根据它们各自的特性采 用不同的分类方法,可将它们进行多种分类,一般采用电泳法和超速离心法进行血浆脂蛋白的分类。 (一)电泳分类法 本法根据不同脂蛋白所带表面电荷不同,在一定外加电场作用下,电泳迁移率不同,可将血浆脂蛋白分为四类。如以硝酸纤维素薄膜为支 持物,电泳结果是:α-脂蛋白泳动最快,相当于α1-球蛋白的位置;前β脂蛋白次之,相当于α2-球蛋白位置;β-脂蛋白泳动在前-β之 后,相当于β-球蛋白的位置;乳糜微粒停留在点样的位置上(见图5-3)。 (二)超速离心法 本法依据不同脂蛋白中蛋白质脂类成分所占比例不同,因而分子密度不同(甘油三酯含量多者密度低,蛋白质含量多的分子密度高),在一 定 离 心 力 作 用 下 , 分 子 沉 降 速 度 或 漂 浮 率 不 同 , 将 脂 蛋 白 分 为 四 类 , 即 乳 糜 微 粒 (chylomicrons) 、 极 低 密 度 脂 蛋 白 (very low densitylipoprotein,VLDL)、低密度脂蛋白(lowdensity lipoprotein,LDL)和高密度脂蛋白(highdensitylipoprotein,HDL);分别相当于电泳分 离中的乳糜微粒、前β脂蛋白、β脂蛋白和α脂蛋白。除上述几类脂蛋白以外,还有一种中间密度脂蛋白(intermediate density lipoprotein,IDL) 其密度位于VLDL与LDL之间,这是VLDL代谢的中间产物。HDL在代谢过程中分子中蛋白与脂类成分有变化,可将HDL再分为HDL1、HDL2 与HDL3。HDL1是在高胆固醇膳食时才出现,HDL2为成熟的HDL,HDL3为新生的HDL,其分子中蛋白成分多。 血浆中的游离中短链脂肪酸可与血浆白蛋白结合而被运输,称之为脂酸白蛋白。由于脂类染色时脂肪酸不着色,所以不易观察,实际上它 的位置与白蛋白相当。 二、血浆脂蛋白的组成 (一)脂蛋白中脂类的组成特点: 除脂酸白蛋白外,各类脂蛋白均含有甘油三酯、磷脂、胆固醇及其酯。但组成比例有很大差异,其中甘油三酯在乳糜微粒中含量为最高, 达其化学组成的90%左右。磷脂含量以HDL为最高,达40%以上。胆固醇及其酯以LDL中最多,几乎占其含量50%。VLDL中以甘油三酯含量 为最多,达60%(见表5-2)。 表5-2 血浆脂蛋白的组成 组成 分类 密度 颗粒直径 S1 电泳 蛋白质 总脂 占总脂的% 合成 功能 (mm) 位置 甘油 胆固 游离 部位 % % 磷脂 胆固醇 三酯 醇 酯 脂肪酸 CM <0.95 90- >400 原点 1-2 98-99 88 8 3 1 小肠粘 转运外源性情 1000 膜细胞 甘油三酯

VLDL 0.95- 30- 20- a2球 7-10 90-93 56 20 15 8 肝细胞 转运内源性 前脂蛋白 1.006 90 400 蛋白 甘油三酯 LDL 1.006 20-25 2-20 球 21 79 13 28 48 10 血浆 转运内源性 脂蛋白 1.063 蛋白 胆固醇 HDL 1.063 12-20 沉降 al球 33 67 16 43 61 10 肝肠 转运外源 a脂蛋白 1.125 蛋白 血浆 性胆因醇 HDL2 1.125 7.5-10 沉降 al球 57 43 13 46 29 6 肝肠 转运内源性 HDL3 1.120 蛋白 血浆 胆固醇 (二)载脂蛋白(apoprotein,apo) 脂蛋白中与脂类结合的蛋白质称为载脂蛋白,载脂蛋白在肝脏和小肠粘膜细胞中合成。目前已发现了十几种载脂蛋白,结构与功能研究比 较清楚的有apoA、apoB、apoC、apoD与apoE五类。每一类脂蛋白又可分为不同的亚类,如apoB分为B100和B48;apoC分为CI、CI、CⅢ 等。载脂蛋白在分子结构上具有一定特点,往往含有较多的双性·螺旋结构,表现出两面性,分子的一侧极性较高可与水溶剂及磷脂或胆固 醇极性区结合,构成脂蛋白的亲水面,分子的另一侧极性较低可与非极性的脂类结合,构成脂蛋白的疏水核心区。 载脂蛋白的主要功能是稳定血浆脂蛋白结构,作为脂类的运输载体。除此以外有些脂蛋白还可作为酶的激活剂:如即oI激活卵磷脂胆固 醇脂酰转移酶(lecithin?cholesteroltransferase,LCAT),apoCⅡ可激活脂蛋白脂肪酶(lipoproteinlipase,LPL)。有些脂蛋白也可作为细胞膜受体的配 体:如apoB-48,apoE参与肝细胞对CM的识别,apoB-100可被各种组织细胞表面LDL受体所识别等(见表5·3). 表5·3血浆脂蛋白的载脂蛋白 载脂蛋白 脂蛋折 氨基酸数 分子量 来源 功能 含量 AI HDL,CM 243 28.300 肝、肠 激活LCAT:HDL受体配基 121.8+13.7 AlI HDL,CM 7 17.500 肝、肠 稳定HDL结构?抑制LCAT? 33.0+5.0* AIV CM,HDL 371 46.000 肠 功能不清(促进CM合成?) 17+2△ B-100 LDL,VLDL,IDL 4536 512,723 肝 LDL受体配基(识别LDL受体) 87.3+143 B-48 CM,CM残粒 265000 肠 促进CM合成 5△ CI VLDL,HDL,CM 57 6,500 肝 激活LCAT? 7.8±2.4* cu VLDL,HDL,CM 79 8,800 肝 激活肝外LPL 5.0±1.8* CⅢ VLDL,HDL,CM 79 8,900 肝 抑制LPL,抑制肝apoE受体 11.8+3.6* 0 HDL 22,100 肝? 功能不清(转达运胆固醇酯? 10±4△ VLDL,HDL,CM LDL受体配基,肝CM残受体) CM残粒 299 34,000 肝 (apoE受体)配基 3.5+12* *华西医科大学生化教研室对625例成都地区正常成人测定的结果。“国外报导参考值。 LCAT:卵磷脂胆固醇脂酰转移酶LPL:脂蛋白脂肪酶 (三)脂蛋白的代谢 1.乳糜微粒(CM) 乳糜微粒是在小肠粘膜细胞中生成的,食物中的脂类在细胞滑面内质网上经再酯化后与粗面内质网上合成的载脂蛋白构成新生的(ascent) 乳糜微粒(包括甘油三酯、胆固醇酯和磷脂以及oB48),经高尔基复合体分泌到细胞外,进入淋巴循环最终进入血液。 新生乳糜微粒入血后,接受来自HDL的apoC和apoE,同时失去部分apoA,被修饰成为成熟的乳糜微粒。成熟分子上的apoCⅡ可激活脂蛋 白脂肪酶(LPL)催化乳糜微粒中甘油三酯水解为甘油和脂肪。此酶存在于脂肪组织、心和肌肉组织的毛细血管内皮细胞外表面上。脂肪酸可被 上述组织摄取而利用,甘油可进入肝脏用于糖异生。通过LPL的作用,乳糜微粒中的甘油三酯大部分被水解利用,同时apoA、apoC、胆固醇 和磷脂转移到HDL上,CM逐渐变小,成为以含胆固醇酯为主的乳糜微粒残余颗粒(remnant)。肝细胞膜上的apoE受体可识别CM残余颗粒,将 其吞噬入肝细胞,与细胞溶酶体融合,载脂蛋白被水解为氨基酸,胆固醇酯分解为胆固醇和脂肪酸,进而可被肝脏利用或分解,完成最终代谢 (图5-4). 黄物中粉类 新生的乳廉微拉 G ch 月APO-E APO-A HDI 组出略弱肪酸 TO,chE 水师 图5.4乳糜微粒(CM)的代谢过程 TG:甘油三酯;pL:磷脂:Ch:胆固醇;ChE:胆固醇酯;ApO:载脂蛋白;HDL:高密度脂蛋白。 由此可见,CM代谢的主要功能就是将外源性甘油三酯转运至脂肪、心和肌肉等肝外组织而利用,同时将食物中外源性胆固醇转运至肝 脏
VLDL 0.95- 30- 20- α2球 7-10 90-93 56 20 15 8 1 肝细胞 转运内源性 前β脂蛋白 1.006 90 400 蛋白 甘油三酯 LDL 1.006- 20-25 2-20 β球 21 79 13 28 48 10 1 血浆 转运内源性 β脂蛋白 1.063 蛋白 胆固醇 HDL 1.063- 12-20 沉降 α1球 33 67 16 43 61 10 4 肝肠 转运外源 α脂蛋白 1.125 蛋白 血浆 性胆固醇 HDL2 1.125 7.5-10 沉降 α1球 57 43 13 46 29 6 6 肝肠 转运内源性 HDL3 1.120 蛋白 血浆 胆固醇 (二)载脂蛋白(apoprotein,apo) 脂蛋白中与脂类结合的蛋白质称为载脂蛋白,载脂蛋白在肝脏和小肠粘膜细胞中合成。目前已发现了十几种载脂蛋白,结构与功能研究比 较清楚的有apoA、apoB、apoC、apoD与apoE五类。每一类脂蛋白又可分为不同的亚类,如apoB分为B100和B48;apoC分为CⅠ、CⅡ、CⅢ 等。载脂蛋白在分子结构上具有一定特点,往往含有较多的双性α-螺旋结构,表现出两面性,分子的一侧极性较高可与水溶剂及磷脂或胆固 醇极性区结合,构成脂蛋白的亲水面,分子的另一侧极性较低可与非极性的脂类结合,构成脂蛋白的疏水核心区。 载脂蛋白的主要功能是稳定血浆脂蛋白结构,作为脂类的运输载体。除此以外有些脂蛋白还可作为酶的激活剂:如apoAI激活卵磷脂胆固 醇脂酰转移酶(lecithin?cholesteroltransferase.LCAT),apoCⅡ可激活脂蛋白脂肪酶(lipoproteinlipase,LPL)。有些脂蛋白也可作为细胞膜受体的配 体:如apo B-48,apoE参与肝细胞对CM的识别,apoB-100可被各种组织细胞表面LDL受体所识别等(见表5-3)。 表5-3 血浆脂蛋白的载脂蛋白 载脂蛋白 脂蛋折 氨基酸数 分子量 来源 功能 含量 AⅠ HDL,CM 243 28.300 肝、肠 激活LCAT;HDL受体配基 121.8±13.7* AⅡ HDL,CM 77 17.500 肝、肠 稳定HDL结构?抑制LCAT? 33.0±5.0* AⅣ CM,HDL 371 46.000 肠 功能不清(促进CM合成?) 17±2△ B-100 LDL,VLDL,IDL 4536 512,723 肝 LDL受体配基(识别LDL受体) 87.3±143* B-48 CM,CM残粒 265 000 肠 促进CM合成 5△ CⅠ VLDL,HDL,CM 57 6,500 肝 激活LCAT? 7.8±2.4* CⅡ VLDL,HDL,CM 79 8,800 肝 激活肝外LPL 5.0±1.8* CⅢ VLDL,HDL,CM 79 8,900 肝 抑制LPL,抑制肝apoE受体 11.8±3.6* D HDL 22,100 肝? 功能不清(转达运胆固醇酯? 10±4△ VLDL,HDL,CM LDL受体配基,肝CM残受体) E CM残粒 299 34,000 肝 (apoE受体)配基 3.5±1.2* *华西医科大学生化教研室对625例成都地区正常成人测定的结果。△国外报导参考值。 LCAT:卵磷脂胆固醇脂酰转移酶 LPL:脂蛋白脂肪酶 (三)脂蛋白的代谢 1.乳糜微粒(CM) 乳糜微粒是在小肠粘膜细胞中生成的,食物中的脂类在细胞滑面内质网上经再酯化后与粗面内质网上合成的载脂蛋白构成新生的(nascent) 乳糜微粒(包括甘油三酯、胆固醇酯和磷脂以及poB48),经高尔基复合体分泌到细胞外,进入淋巴循环最终进入血液。 新生乳糜微粒入血后,接受来自HDL的apoC和apoE,同时失去部分apoA,被修饰成为成熟的乳糜微粒。成熟分子上的apoCⅡ可激活脂蛋 白脂肪酶(LPL)催化乳糜微粒中甘油三酯水解为甘油和脂肪。此酶存在于脂肪组织、心和肌肉组织的毛细血管内皮细胞外表面上。脂肪酸可被 上述组织摄取而利用,甘油可进入肝脏用于糖异生。通过LPL的作用,乳糜微粒中的甘油三酯大部分被水解利用,同时apoA、apoC、胆固醇 和磷脂转移到HDL上,CM逐渐变小,成为以含胆固醇酯为主的乳糜微粒残余颗粒(remnant)。肝细胞膜上的apoE受体可识别CM残余颗粒,将 其吞噬入肝细胞,与细胞溶酶体融合,载脂蛋白被水解为氨基酸,胆固醇酯分解为胆固醇和脂肪酸,进而可被肝脏利用或分解,完成最终代谢 (图5-4)。 图5-4 乳糜微粒(CM)的代谢过程 TG:甘油三酯;pL:磷脂;Ch:胆固醇;ChE:胆固醇酯;ApO:载脂蛋白;HDL:高密度脂蛋白。 由此可见,CM代谢的主要功能就是将外源性甘油三酯转运至脂肪、心和肌肉等肝外组织而利用,同时将食物中外源性胆固醇转运至肝 脏
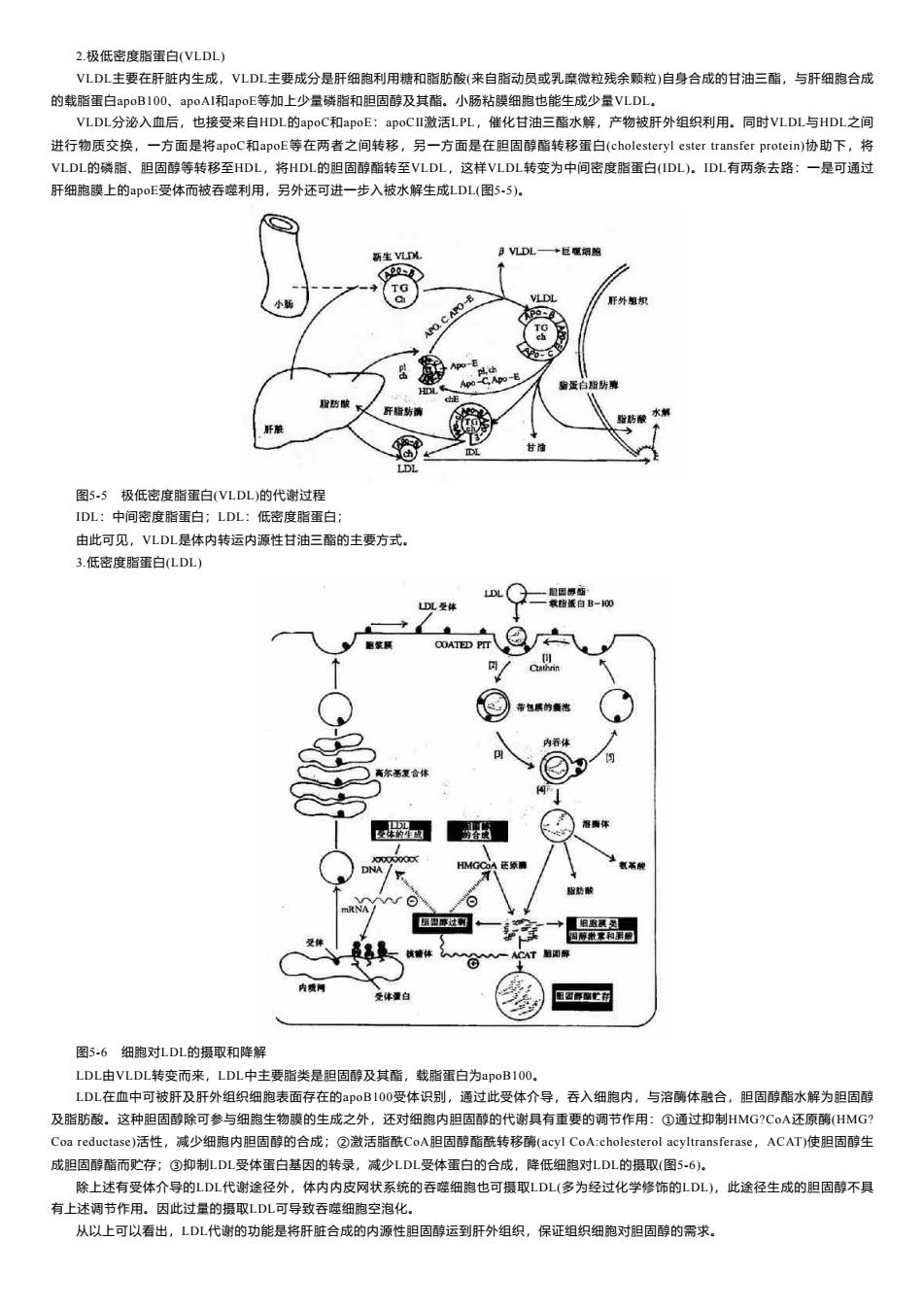
2.极低密度脂蛋白(VLDL) VLDL主要在肝脏内生成,VLDL主要成分是肝细胞利用糖和脂肪酸(来自脂动员或乳糜微粒残余颗粒)自身合成的甘油三酯,与肝细胞合成 的载脂蛋白apoB100、apoAli和apoE等加上少量磷脂和胆固醇及其酯。小肠粘膜细胞也能生成少量VLDL。 VLDL分泌入血后,也接受来自HDL的apoC和apoE:apoCⅡ激活LPL,催化甘油三酯水解,产物被肝外组织利用。同时VLDL与HDL之间 进行物质交换,一方面是将apoC和apoE等在两者之间转移,另一方面是在胆固醇酯转移蛋白(cholesteryl ester transfer protein)协助下,将 VLDL的磷脂、胆固醇等转移至HDL,将HDL的胆固醇酯转至VLDL,这样VLDL转变为中间密度脂蛋白(IDL)。IDL有两条去路:一是可通过 肝细胞膜上的apoE受体而被吞噬利用,另外还可进一步入被水解生成LDL(图5-5)。 新生VLD以 月V儿DL+巨赚细购 小 CAPO-E VLDL 肝外组织 APO Apo-E HDL黑 贴蛋 程防酸 肝后防膜 肝胜 甘油 LDL 图5-5极低密度脂蛋白(VLDL)的代谢过程 IDL:中间密度脂蛋白:LDL:低密度脂蛋白: 由此可见,VLDL是体内转运内源性甘油三酯的主要方式。 3.低密度脂蛋白(LDL) LDL DL受体 载脂蛋白B-00 脑展 COATED Pr 酸 酸药酸 团型摩过网 细虫顶卖 一ACAT自圆醇 体量白 图5-6细胞对LDL的摄取和降解 LDL由VLDL转变而来,LDL中主要脂类是胆固醇及其酯,载脂蛋白为apoB1O0。 LDL在血中可被肝及肝外组织细胞表面存在的oB100受体识别,通过此受体介导,吞入细胞内,与溶酶体融合,胆固醇酯水解为胆固醇 及脂肪酸。这种胆固醇除可参与细胞生物膜的生成之外,还对细胞内胆固醇的代谢具有重要的调节作用:①通过抑制HMG?CoA还原酶(HMG? Coa reductase)活性,减少细胞内胆固醇的合成;②激活脂酰CoA胆固醇酯酰转移酶(acyl CoA:cholesterol acyltransferase,ACAT)使胆固醇生 成胆固醇酯而贮存;③抑制LDL受体蛋白基因的转录,减少LDL受体蛋白的合成,降低细胞对LDL的摄取(图5-6). 除上述有受体介导的LDL代谢途径外,体内内皮网状系统的吞噬细胞也可摄取LDL(多为经过化学修饰的LDL),此途径生成的胆固醇不具 有上述调节作用。因此过量的摄取LDL可导致吞噬细胞空泡化。 从以上可以看出,LDL代谢的功能是将肝脏合成的内源性胆固醇运到肝外组织,保证组织细胞对胆固醇的需求
2.极低密度脂蛋白(VLDL) VLDL主要在肝脏内生成,VLDL主要成分是肝细胞利用糖和脂肪酸(来自脂动员或乳糜微粒残余颗粒)自身合成的甘油三酯,与肝细胞合成 的载脂蛋白apoB100、apoAI和apoE等加上少量磷脂和胆固醇及其酯。小肠粘膜细胞也能生成少量VLDL。 VLDL分泌入血后,也接受来自HDL的apoC和apoE:apoCⅡ激活LPL,催化甘油三酯水解,产物被肝外组织利用。同时VLDL与HDL之间 进行物质交换,一方面是将apoC和apoE等在两者之间转移,另一方面是在胆固醇酯转移蛋白(cholesteryl ester transfer protein)协助下,将 VLDL的磷脂、胆固醇等转移至HDL,将HDL的胆固醇酯转至VLDL,这样VLDL转变为中间密度脂蛋白(IDL)。IDL有两条去路:一是可通过 肝细胞膜上的apoE受体而被吞噬利用,另外还可进一步入被水解生成LDL(图5-5)。 图5-5 极低密度脂蛋白(VLDL)的代谢过程 IDL:中间密度脂蛋白;LDL:低密度脂蛋白; 由此可见,VLDL是体内转运内源性甘油三酯的主要方式。 3.低密度脂蛋白(LDL) 图5-6 细胞对LDL的摄取和降解 LDL由VLDL转变而来,LDL中主要脂类是胆固醇及其酯,载脂蛋白为apoB100。 LDL在血中可被肝及肝外组织细胞表面存在的apoB100受体识别,通过此受体介导,吞入细胞内,与溶酶体融合,胆固醇酯水解为胆固醇 及脂肪酸。这种胆固醇除可参与细胞生物膜的生成之外,还对细胞内胆固醇的代谢具有重要的调节作用:①通过抑制HMG?CoA还原酶(HMG? Coa reductase)活性,减少细胞内胆固醇的合成;②激活脂酰CoA胆固醇酯酰转移酶(acyl CoA:cholesterol acyltransferase,ACAT)使胆固醇生 成胆固醇酯而贮存;③抑制LDL受体蛋白基因的转录,减少LDL受体蛋白的合成,降低细胞对LDL的摄取(图5-6)。 除上述有受体介导的LDL代谢途径外,体内内皮网状系统的吞噬细胞也可摄取LDL(多为经过化学修饰的LDL),此途径生成的胆固醇不具 有上述调节作用。因此过量的摄取LDL可导致吞噬细胞空泡化。 从以上可以看出,LDL代谢的功能是将肝脏合成的内源性胆固醇运到肝外组织,保证组织细胞对胆固醇的需求