
第四章糖代谢 糖是有机体重要的能源和碳源。糖代谢包括糖的合成与糖的分解两方面。糖的最终来 源都是植物或光合细菌通过光合作用将CO和水同化成葡萄糖。除此之外糖的合成途径还 包括糖的异生一非糖物质转化成糖的途径。在植物和动物体内葡萄糖可以进一步合成寡糖 和多糖作为储能物质(如蔗糖、淀粉和糖元),或者构成植物或细菌的细胞壁(如纤维素和肽聚 糖)。 在生物体内,糖(主要是葡萄糖)的降解是生命活动所需能如ATP)的来源。生物体从 碳水化合物中获得能量大致分成三个阶段:在第一阶段,大分子糖变成小分子糖,如淀粉 糖元等变成葡萄糖:在第二阶段,葡萄糖通过糖酵解(糖的共同分解途径)降解为丙酮酸, 丙酮酸再转变为活化的酰基载体一乙酰辅酶A:在第三阶段,乙酰辅酶A通过三羧酸循环 (糖的最后氧化途径)彻底氧化成CO,当电子传递给最终的电子受体O时生成ATP。这是 动物、植物和微生物获得能量以维持生存的共同途径。糖的中间代谢还包括磷酸戊糖途径、 乙醛酸途径等。 第一节新陈代谢慨论 一、新陈代谢的意义 新陈代谢是生物最基本的特征之一。新陈代谢是指生物活体与外界环境不断交换物质 的过程。机体从外界摄取营养物质,转化为机体自身需要的物质称为同化作用.是由小分 子合成生物大分子,需要能量:而机体自身原有的物质的分解、排泄称为异化作用,是由 生物大分子降解为生物小分子,最后分解成C0和出0,释放能量。异化作用释放的能量 可供机体生理活动的需要。同化作用为异化作用提供物质基础,异化作用为同化作用提供 能量基础。同化作用和异化作用是既对立又统一的矛盾的两个方面,两者相互联系、相互 制约,互为基础。 生物机体的同化作用和异化作用都包含着一系列逐步进行的合成与分解的反应,称之 为中间代谢反应。能量的释放与供应是逐步进行的,也是由许多中间代谢反应组成的。 机体与外界环境进行物质交换的过程称为物质代谢。在物质交换中伴随着能量的交 换,也称为能量代谢。植物通过光合作用将太阳的光能转变为糖的化学能。当糖在体内进 行分解代谢时,再将化学能释放出来,用于合成代谢,也可转变为机械能、光能、电能等 各种形式的能,以满足生命活动的需要。同化作用和异化作用、物质代谢和能量代谢的相 互关系可用图解表示如下: 合成代谢「生物小分子合成为生物大分子 (同化作用)1需要能量1 新陈代谢 能量代谢 物质代谢 释放能量 分解代谢 (异化作用)【生物大分子分解为生物小分子 143
143 第四章 糖代谢 糖是有机体重要的能源和碳源。糖代谢包括糖的合成与糖的分解两方面。糖的最终来 源都是植物或光合细菌通过光合作用将 CO2和水同化成葡萄糖。除此之外糖的合成途径还 包括糖的异生—非糖物质转化成糖的途径。在植物和动物体内葡萄糖可以进一步合成寡糖 和多糖作为储能物质(如蔗糖、淀粉和糖元),或者构成植物或细菌的细胞壁(如纤维素和肽聚 糖)。 在生物体内,糖(主要是葡萄糖)的降解是生命活动所需能量(如 ATP)的来源。生物体从 碳水化合物中获得能量大致分成三个阶段:在第一阶段,大分子糖变成小分子糖,如淀粉、 糖元等变成葡萄糖;在第二阶段,葡萄糖通过糖酵解(糖的共同分解途径)降解为丙酮酸, 丙酮酸再转变为活化的酰基载体—乙酰辅酶 A;在第三阶段,乙酰辅酶 A 通过三羧酸循环 (糖的最后氧化途径)彻底氧化成 CO2,当电子传递给最终的电子受体 O2时生成 ATP。这是 动物、植物和微生物获得能量以维持生存的共同途径。糖的中间代谢还包括磷酸戊糖途径、 乙醛酸途径等。 第一节 新陈代谢概论 一、新陈代谢的意义 新陈代谢是生物最基本的特征之一。新陈代谢是指生物活体与外界环境不断交换物质 的过程。机体从外界摄取营养物质,转化为机体自身需要的物质称为同化作用,是由小分 子合成生物大分子,需要能量;而机体自身原有的物质的分解、排泄称为异化作用,是由 生物大分子降解为生物小分子,最后分解成 CO2 和 H2O,释放能量。异化作用释放的能量 可供机体生理活动的需要。同化作用为异化作用提供物质基础,异化作用为同化作用提供 能量基础。同化作用和异化作用是既对立又统一的矛盾的两个方面,两者相互联系、相互 制约,互为基础。 生物机体的同化作用和异化作用都包含着一系列逐步进行的合成与分解的反应,称之 为中间代谢反应。能量的释放与供应是逐步进行的,也是由许多中间代谢反应组成的。 机体与外界环境进行物质交换的过程称为物质代谢。在物质交换中伴随着能量的交 换,也称为能量代谢。植物通过光合作用将太阳的光能转变为糖的化学能。当糖在体内进 行分解代谢时,再将化学能释放出来,用于合成代谢,也可转变为机械能、光能、电能等 各种形式的能,以满足生命活动的需要。同化作用和异化作用、物质代谢和能量代谢的相 互关系可用图解表示如下: 新陈代谢 合成代谢 (同化作用) 分解代谢 (异化作用) 生物小分子合成为生物大分子 需要能量 释放能量 能量代谢 生物大分子分解为生物小分子 物质代谢

各种生物的新陈代谢过程虽然复 脂防 多 蛋白质 杂,却有共同的特点:绝大多数 新陈代谢反应是在温和的条件下 一阶段 复基酸 进行的:新陈代谢的一系列中间 脂斯酸及甘油葡萄及其它籍 反应相互配合、有条不系、彼此 一阶到 协调,又具有严格的倾序性:三 大营养物质(糖、脂类、蛋白质) 的分解且右共同途径,二者均出 转化为丙酮酸,再经三羧酸循环 氧化成C0和水(见图61):具有 氧化 高度的适应性和灵敏性的自动调 节控制体系以适应内外条件的变 化。 图61生物获得能量的三个阶段 第二节双糖和多糖的酶促降解 一、蔗糖、麦芽糖和乳糖的酶促降解 (I)蔗糖的水解蔗糖的水解主要通过转化酶的作用,转化酶又称蔗糖酶(sucras©),它 广泛存在于植物体内。所有的转化酶都是B-果糖苷酶。其作用如下: 以上反应是不可逆的,因为蔗糖水解时放出大量的热能 (2)乳糖的水解乳糖的水解由乳糖酶(又称为B半乳糖苷酶)催化,生成半乳糖和葡 萄糖: 乳糖+H0 乳糖酶 半乳糖十葡萄糖 (3)麦芽糖的水解由麦芽糖酶催化1分子麦芽糖水解生成2分子葡萄糖: 麦芽糖+0麦珍糖酶 一2萄萄糖 二、淀粉(糖元)的酶促降解 淀粉可以通过两种不同的过程降解成葡萄糖。一个过程是水解,动物的消化或植物种 子萌发时就是利用这一途径使多糖降解成糊精、麦芽糖、异麦芽糖和葡萄糖。其中的麦芽 糖和异麦芽糖又可被麦芽糖酶和异麦芽糖酶降解生成葡萄糖。葡萄糖进入细胞后被磷酸化 并经糖酵解作用降解。淀粉的另一个降解途径为磷酸降解过程。 1.淀粉的水解催化淀粉水解的酶称为淀粉酶(amylase)它又可分两种:一种称为 144
144 各种生物的新陈代谢过程虽然复 杂,却有共同的特点:绝大多数 新陈代谢反应是在温和的条件下 进行的;新陈代谢的一系列中间 反应相互配合、有条不紊、彼此 协调,又具有严格的顺序性;三 大营养物质(糖、脂类、蛋白质) 的分解具有共同途径,三者均先 转化为丙酮酸,再经三羧酸循环 氧化成 CO2和水(见图 6-1);具有 高度的适应性和灵敏性的自动调 节控制体系以适应内外条件的变 化。 图 6-1 生物获得能量的三个阶段 第二节 双糖和多糖的酶促降解 一、蔗糖、麦芽糖和乳糖的酶促降解 (1)蔗糖的水解 蔗糖的水解主要通过转化酶的作用,转化酶又称蔗糖酶(sucrase),它 广泛存在于植物体内。所有的转化酶都是β-果糖苷酶。其作用如下: 转化酶 蔗糖+H2O────→ 葡萄糖+果糖 以上反应是不可逆的,因为蔗糖水解时放出大量的热能。 (2)乳糖的水解 乳糖的水解由乳糖酶(又称为β-半乳糖苷酶)催化,生成半乳糖和葡 萄糖: 乳糖酶 乳糖+H2O ────→ 半乳糖+葡萄糖 (3)麦芽糖的水解 由麦芽糖酶催化 1 分子麦芽糖水解生成 2 分子葡萄糖: 麦芽糖酶 麦芽糖+H2O ────→ 2 葡萄糖 二、淀粉(糖元)的酶促降解 淀粉可以通过两种不同的过程降解成葡萄糖。一个过程是水解,动物的消化或植物种 子萌发时就是利用这一途径使多糖降解成糊精、麦芽糖、异麦芽糖和葡萄糖。其中的麦芽 糖和异麦芽糖又可被麦芽糖酶和异麦芽糖酶降解生成葡萄糖。葡萄糖进入细胞后被磷酸化 并经糖酵解作用降解。淀粉的另一个降解途径为磷酸降解过程。 l.淀粉的水解 催化淀粉水解的酶称为淀粉酶(amylase)它又可分两种:一种称为

a-l,4葡聚糖水解酶,又称为a-淀粉酶(a-amylase)),是一种内淀粉酶endoamy-ase),能 以一种无规则的方式水解直链淀粉amylose)内部的锭,生成葡萄糖与麦芽糖的混合物,如 果底物是支链淀粉(amylopectin),则水解产物中含有支链和非支链的寡聚糖类的混合物, 其中存在āL6键。第二种水解酶称为α-1,4葡聚糖基麦芽糖基水解酶,又称为B-淀粉 酶,是一种外淀粉酶(exoamylase)©,它作用于多糖的非还原性末端而生成麦芽糖,所以当B -淀粉酶作用于直链淀粉时能生成定量的麦芽糖。当底物为分支的支链淀粉或糖元时,则生 成的产物为麦芽糖和多分枝糊精,因为此酶仅能作用于ā山,4键而不能作用于α-6键。淀 粉酶在动物、植物及微生物中均存在。在动物中主要在消化液(唾液及胰液)中存在。图6-2 为α·淀粉酶及B-淀粉酶水解支链淀粉的示意图。 ā-淀粉酶仅在发芽的种子 0D-0①-①合号台 中存在,如大麦发芽后,则a 淀粉酶及B-淀粉酶均有存在。在 ●-●●● pH值3.3时,a-淀粉酶就被破 ①0-0-●●●- 坏,但它能耐高温,温度高达 数个1情酸有有 70℃(约15min)仍稳定。而B. 淀粉酶主要存在于休眠的种子 ee-o 中,在0C高温时容易破坏,但 对酸比较稳定,在pH值3.3时 ッ 仍不被破坏,所以利用高温或调 转移酶 节pH值的方法可以将这两种淀 粉酶分开。这两种淀粉酶现在都 能制成结晶。α-淀粉酶和B淀 粉酶中的ā与B,并非表示其作 999y2: 用于a或B糖苷键,而只是用米 1一1,6轴苷酵 标明两种不同的水解淀粉的酶 由于ā-淀粉酶和B-淀粉酶只能 水解淀粉的ā-1.4鞋,因此只能 999, 使支链淀粉水解54%~55%,剩 下的分支组成了一个淀粉酶不能 图63糖原降解步骤 作用的糊精,称为极限糊精 极限糊精中的ā1,6键可被R酶水解,R酶又称脱支酶,脱支酶仅能分解支链淀粉外 围的分支,不能分解支链淀粉内部的分支,只有与B淀粉酶共同作用才能将支链淀粉完全 降解,生成麦芽糖和葡萄糖。麦芽糖被麦芽糖酶水解生成葡萄糖,进一步被植物利用。 2.淀粉的磷酸解 ①)ā-l,4键的降解淀粉的磷酸解是在淀粉磷酸化酶的催化下,用磷酸代替水,将淀 粉降解生成磷酸葡萄糖的作用。 淀粉磷酸解的好处是:生成的产物上磷酸葡萄糖不能扩散到细胞外,可直接进入糖酵 解途径,节省了能量:而淀粉的水解产物葡萄糖,能进行扩散,但还必须经过磷酸化消耗 145
145 α-1,4-葡聚糖水解酶,又称为α-淀粉酶(α-amylase),是一种内淀粉酶(endoamy-lase),能 以一种无规则的方式水解直链淀粉(amylose)内部的键,生成葡萄糖与麦芽糖的混合物,如 果底物是支链淀粉(amylopectin),则水解产物中含有支链和非支链的寡聚糖类的混合物, 其中存在α-l,6 键。第二种水解酶称为 α-1,4-葡聚糖基-麦芽糖基水解酶,又称为β-淀粉 酶,是一种外淀粉酶(exoamylase),它作用于多糖的非还原性末端而生成麦芽糖,所以当β -淀粉酶作用于直链淀粉时能生成定量的麦芽糖。当底物为分支的支链淀粉或糖元时,则生 成的产物为麦芽糖和多分枝糊精,因为此酶仅能作用于α-l,4键而不能作用于α-l,6 键。淀 粉酶在动物、植物及微生物中均存在。在动物中主要在消化液(唾液及胰液)中存在。图 6-2 为α-淀粉酶及β-淀粉酶水解支链淀粉的示意图。 α-淀粉酶仅在发芽的种子 中存在,如大麦发芽后,则α- 淀粉酶及β-淀粉酶均有存在。在 pH 值 3.3 时,α-淀粉酶就被破 坏,但它能耐高温,温度高达 70℃(约 15 min)仍稳定 。而β- 淀粉酶主要存在于休眠的种子 中,在 70℃高温时容易破坏,但 对酸比较稳定,在 pH 值 3.3 时 仍不被破坏,所以利用高温或调 节 pH 值的方法可以将这两种淀 粉酶分开。这两种淀粉酶现在都 能制成结晶。α-淀粉酶和β-淀 粉酶中的α与β,并非表示其作 用于α或β糖苷键,而只是用来 标明两种不同的水解淀粉的酶。 由于α-淀粉酶和β-淀粉酶只能 水解淀粉的α-1,4 键,因此只能 使支链淀粉水解 54%~55%,剩 下的分支组成了一个淀粉酶不能 作用的糊精,称为极限糊精。 极限糊精中的α-1,6 键可被 R 酶水解,R 酶又称脱支酶,脱支酶仅能分解支链淀粉外 围的分支,不能分解支链淀粉内部的分支,只有与β-淀粉酶共同作用才能将支链淀粉完全 降解,生成麦芽糖和葡萄糖。麦芽糖被麦芽糖酶水解生成葡萄糖,进一步被植物利用。 2.淀粉的磷酸解 (l)α-l,4 键的降解 淀粉的磷酸解是在淀粉磷酸化酶的催化下,用磷酸代替水,将淀 粉降解生成 l-磷酸葡萄糖的作用。 淀粉磷酸解的好处是:生成的产物 l-磷酸葡萄糖不能扩散到细胞外,可直接进入糖酵 解途径,节省了能量;而淀粉的水解产物葡萄糖,能进行扩散,但还必须经过磷酸化消耗 图 6-3 糖原降解步骤

一个ATP才能进入糖酵解途径。 淀粉磷酸解步骤可表示如下: 淀粉 !淀粉碳酸化酷 1-磷酸葡萄糖 !磷酸萄档越变位码 6-磷酸葡萄糖 !6陵酸萄苟糖脂面 葡萄糖+Pi (2)α1,6支链的降解α-淀粉酶、B-淀粉酶和淀粉磷酸化酶只能水解淀粉(或糖原)的 a-1,4键,不能水解a-1,6键。由图63可见,磷酸化酶将一个分支上的5个αL,4糖苷键 和另一个分支上的3个ā-」4轴件罐水解,至末端残基a和d处即停止,出时需要一个转 移酶,将一个分支上的3个糖残基(abc)转移到另一个分支上,在糖残基c与d之间形成 个新的a-1.4键,然后在a-L6糖苷酶的作用下,水解z与h之间的a-1.6糖苷键,从而将 个具有分支结构的糖原转变成为线型的直链结构,后者可被磷酸化酶继续分解。因此, 淀粉(或糖原)降解生成葡萄糖是几种酶相互配合进行催化反应的结果。 三、细胞壁多糖的酶促降解 1.纤维素的降解纤维素酶可使纤维素分子的B1,4键发生水解,生成纤维二糖,在 纤维二糖酶的作用下,最后分解为B-葡萄糖,反应过程如下: 纤维素酶 纤维素 一纤维二郁年霜二糖酚 一B萄萄糖 纤维素的分解在高等植物体内很少发生,只是在少数发芽的种子及其幼苗(如大麦、 菠菜、玉米等内发现有纤维素酶的分解作用。但在许多微生物体内(如细菌、霉菌)都含有 分解纤维素的酶。 2.果胶的降解果胶酶是植物体中催化果胶物质水解的酶。果胶酶按其所水解的键, 可分两种:一种称果胶甲酯酶[(pectinesterase)(PE)或果胶酶:另一种是半乳糖醛酸酶 [(Polygalacturonase)(CPG)。果胶甲脂酶水解果胶酸的甲酯,生成果胶酸和甲醇: 果胶甲酯酶 果胶一 →果胶酸十甲醇 半乳糖醛酸酶水解聚半乳糖醛酸之间的α1,4糖苷键,生成半乳糖醛酸。 植物体内一些生理现象与果胶酶的作用有关,如叶柄离层的形成就是果胶酶分解胞间 层的果胶质使细胞相互分离以致叶片脱落:果实成熟时,由于果胶酶的作用使果肉细胞分 离,果肉变软植物感病后,病原菌分泌果胶酶将寄主细胞分离而侵入植物体内。 146
146 一个 ATP 才能进入糖酵解途径。 淀粉磷酸解步骤可表示如下: 淀粉 ↓淀粉磷酸化酶 1-磷酸葡萄糖 ↓磷酸葡萄糖变位酶 6-磷酸葡萄糖 ↓6-磷酸葡萄糖脂酶 葡萄糖+Pi (2)α-l,6支链的降解 α-淀粉酶、β-淀粉酶和淀粉磷酸化酶只能水解淀粉(或糖原)的 α-1,4 键,不能水解α-1,6键。由图 6-3 可见,磷酸化酶将一个分支上的 5 个α-l,4糖苷键 和另一个分支上的 3 个α-l,4 糖苷键水解,至末端残基 a 和 d 处即停止,此时需要一个转 移酶,将一个分支上的 3 个糖残基(abc)转移到另一个分支上,在糖残基 c 与 d 之间形成一 个新的α-1,4 键,然后在α-l,6 糖苷酶的作用下,水解 z 与 h 之间的α-1,6 糖苷键,从而将 一个具有分支结构的糖原转变成为线型的直链结构,后者可被磷酸化酶继续分解。因此, 淀粉(或糖原)降解生成葡萄糖是几种酶相互配合进行催化反应的结果。 三、细胞壁多糖的酶促降解 1.纤维素的降解 纤维素酶可使纤维素分子的β-1,4键发生水解,生成纤维二糖,在 纤维二糖酶的作用下,最后分解为β-葡萄糖,反应过程如下: 纤维素酶 纤维二糖酶 纤维素──────→纤维二糖───────→β-葡萄糖 纤维素的分解在高等植物体内很少发生,只是在少数发芽的种子及其幼苗(如大麦、 菠菜、玉米等)内发现有纤维素酶的分解作用。但在许多微生物体内(如细菌、霉菌)都含有 分解纤维素的酶。 2.果胶的降解 果胶酶是植物体中催化果胶物质水解的酶。果胶酶按其所水解的键, 可分两种:一种称果胶甲酯酶[(pectinesterase) (PE)]或果胶酶;另一种是半乳糖醛酸酶 [(Polygalacturonase) (CPG)]。果胶甲脂酶水解果胶酸的甲酯,生成果胶酸和甲醇: 果胶甲酯酶 果胶───────→果胶酸十甲醇 半乳糖醛酸酶水解聚半乳糖醛酸之间的α-1,4 糖苷键,生成半乳糖醛酸。 植物体内一些生理现象与果胶酶的作用有关,如叶柄离层的形成就是果胶酶分解胞间 层的果胶质使细胞相互分离以致叶片脱落;果实成熟时,由于果胶酶的作用使果肉细胞分 离,果肉变软;植物感病后,病原菌分泌果胶酶将寄主细胞分离而侵入植物体内
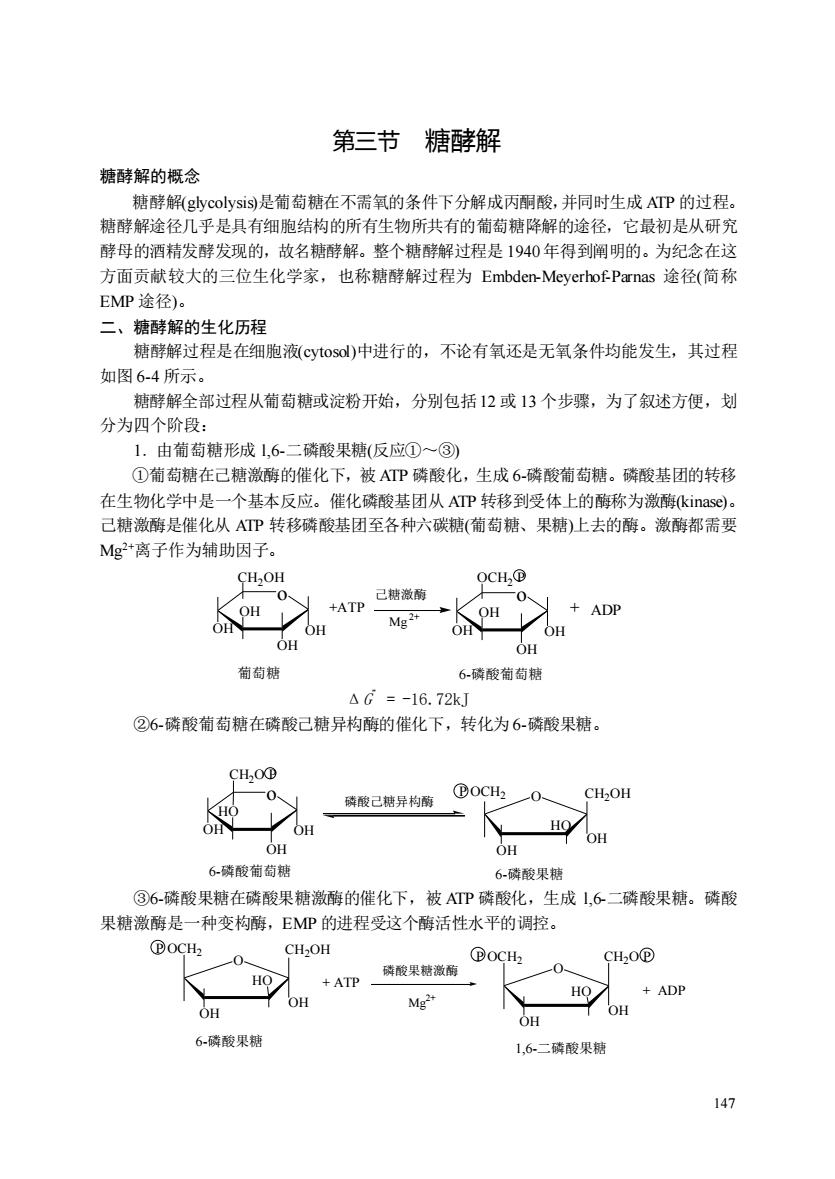
第三节糖酵解 糖酵解的概念 糖酵解(glycolysis9是葡萄糖在不需氧的条件下分解成丙酮酸,并同时生成ATP的过程。 糖酵解途径几乎是具有细胞结构的所有生物所共有的葡萄糖降解的途径,它最初是从研究 酵母的酒精发酵发现的,故名糖酵解。整个糖酵解过程是1940年得到阐明的。为纪念在这 方面贡献较大的三位生化学家,也称糖酵解过程为Embden-Meyerhof-Parnas途径(简称 EMP途径). 二、糖酵解的生化历程 糖酵解过程是在细胞液cytosol)中进行的,不论有氧还是无氧条件均能发生,其过程 如图6-4所示。 糖酵解全部过程从葡萄糖或淀粉开始,分别包括12或13个步骤,为了叙述方便,划 分为四个阶段: 1.由箱萄糖形成1.6-二磷酸果糖(反①)~一③》 ①葡萄糖在己糖激酶的催化下,被ATP磷酸化,生成6-磷酸葡萄糖。磷酸基团的转移 在生物化学中是一个基本反应。催化磷酸基团从ATP转移到受体上的酶称为激酶kinas©)。 己糖激酶是催化从ATP转移磷酸基团至各种六碳糖(葡萄糖、果糖)上去的酶。激酶都需要 Mg*离子作为辅助因子。 OCH.B +ADP OHOH OH OH OH 萄萄糖 6磷酸葡萄糖 △G=-16.72kJ ②6-磷酸葡萄糖在磷酸己糖异构酶的催化下,转化为6磷酸果糖。 CHOD 磷酸己糖异枸箭®0C刊一0 CH2OH OH HO OH OH 6-磷酸葡萄糖 6-磷酸果糖 ③6-磷酸果糖在磷酸果糖激酶的催化下,被ATP磷酸化,生成1,6二磷酸果糖。磷酸 果糖激酶是一种变构酶,EMP的进程受这个酶活性水平的调控。 0CH CH2OH +ATP磷酸果糖 oCHh 0 CH,0@ HO OH HO OH OH 6-磷酸果糖 1,6二磷酸果糖 147
147 第三节 糖酵解 糖酵解的概念 糖酵解(glycolysis)是葡萄糖在不需氧的条件下分解成丙酮酸,并同时生成 ATP 的过程。 糖酵解途径几乎是具有细胞结构的所有生物所共有的葡萄糖降解的途径,它最初是从研究 酵母的酒精发酵发现的,故名糖酵解。整个糖酵解过程是 1940年得到阐明的。为纪念在这 方面贡献较大的三位生化学家,也称糖酵解过程为 Embden-Meyerhof-Parnas 途径(简称 EMP 途径)。 二、糖酵解的生化历程 糖酵解过程是在细胞液(cytosol)中进行的,不论有氧还是无氧条件均能发生,其过程 如图 6-4 所示。 糖酵解全部过程从葡萄糖或淀粉开始,分别包括12 或 13 个步骤,为了叙述方便,划 分为四个阶段: 1.由葡萄糖形成 l,6-二磷酸果糖(反应①~③) ①葡萄糖在己糖激酶的催化下,被 ATP 磷酸化,生成 6-磷酸葡萄糖。磷酸基团的转移 在生物化学中是一个基本反应。催化磷酸基团从 ATP 转移到受体上的酶称为激酶(kinase)。 己糖激酶是催化从 ATP 转移磷酸基团至各种六碳糖(葡萄糖、果糖)上去的酶。激酶都需要 Mg2+离子作为辅助因子。 o OH OH OH OH CH2OH Mg 2+ o OH OH OH OH OCH2 P +ATP + ADP 己糖激酶 葡萄糖 6-磷酸葡萄糖 ΔG ” = -16.72kJ ②6-磷酸葡萄糖在磷酸己糖异构酶的催化下,转化为 6-磷酸果糖。 o OH OH OH HO CH2O P 6-磷酸葡萄糖 磷酸己糖异构酶 OH HO O CH2OH OH 6-磷酸果糖 P OCH2 ③6-磷酸果糖在磷酸果糖激酶的催化下,被 ATP 磷酸化,生成 l,6-二磷酸果糖。磷酸 果糖激酶是一种变构酶,EMP 的进程受这个酶活性水平的调控。 OH HO O CH2OH OH 6-磷酸果糖 P OCH2 OH HO O CH2O P OH 1,6-二磷酸果糖 P OCH2 + ATP 磷酸果糖激酶 Mg 2+ + ADP