
第十二章 核酸的生物合成 核酸是贮存和传递遗传信息的生物大分子。生物体的遗传信息是以密码的形式编码在 DNA分子上,表现为特定的核苷酸排列顺序。在细胞分裂过程中通过DNA的复制把遗传 信息由亲代传递给子代,在子代的个体发育过程中遗传信息由DNA传递到RNA,最后翻 译成特异的蛋白质,表现出与亲代相似的遗传性状。 在某些情况下RNA也是重要的遗传 物质,如在RNA病毒中RNA具有自我复制的能力,并同时作为mRNA,指导病毒蛋白质 的生物合成。在致癌RNA病毒中,RNA还以逆转录的方式将遗传信息传递给DNA分子。 上述遗传信息的流向称为中心法则(central dogma),它是由F.Crick在1958年最早提出的, 其后又得到不断的补充和完善。 中心法则可简洁的用图11-1表示: DNA 翻译 蛋白质 复制病毒] 图111中心法则简图 图中复制就是指以原来分子为模板,合成出相同分子的过程:转录(或逆转录)就是在 DNA(或RNA)分子上合成出与其核苷酸顺序相对应的RNA(或DNA)的过程:翻译就是在 以rRNA和蛋白质组成的核糖核蛋白体(简称核糖体)上,以mRNA为模板,根据每三个相 邻核苷酸决定一种氨基酸的三联体密码规则,由RNA运送氨基酸,合成出具有特定氨基 酸顺序的蛋白质肽链的过程。 第一节 DNA的生物合成 一、半保留复制 DNA呈双股螺旋结构,这样的结构对于维持遗传物质的稳定性和复制的准确性都是极 270
270 第十二章 核酸的生物合成 核酸是贮存和传递遗传信息的生物大分子。生物体的遗传信息是以密码的形式编码在 DNA 分子上,表现为特定的核苷酸排列顺序。在细胞分裂过程中通过 DNA 的复制把遗传 信息由亲代传递给子代,在子代的个体发育过程中遗传信息由DNA 传递到 RNA,最后翻 译成特异的蛋白质,表现出与亲代相似的遗传性状。在某些情况下 RNA 也是重要的遗传 物质,如在 RNA 病毒中 RNA 具有自我复制的能力,并同时作为 mRNA,指导病毒蛋白质 的生物合成。在致癌 RNA 病毒中,RNA还以逆转录的方式将遗传信息传递给 DNA 分子。 上述遗传信息的流向称为中心法则(central dogma),它是由 F.Crick 在 1958 年最早提出的, 其后又得到不断的补充和完善。 中心法则可简洁的用图 11-1 表示: 图 11-1 中心法则简图 图中复制就是指以原来分子为模板,合成出相同分子的过程;转录(或逆转录)就是在 DNA(或 RNA)分子上合成出与其核苷酸顺序相对应的 RNA(或 DNA)的过程;翻译就是在 以 rRNA 和蛋白质组成的核糖核蛋白体(简称核糖体)上,以 mRNA 为模板,根据每三个相 邻核苷酸决定一种氨基酸的三联体密码规则,由 tRNA 运送氨基酸,合成出具有特定氨基 酸顺序的蛋白质肽链的过程。 第一节 DNA 的生物合成 一、半保留复制 DNA 呈双股螺旋结构,这样的结构对于维持遗传物质的稳定性和复制的准确性都是极 DNA 复制 转录 逆转录 复制 [病毒] 翻译 蛋白质 RNA
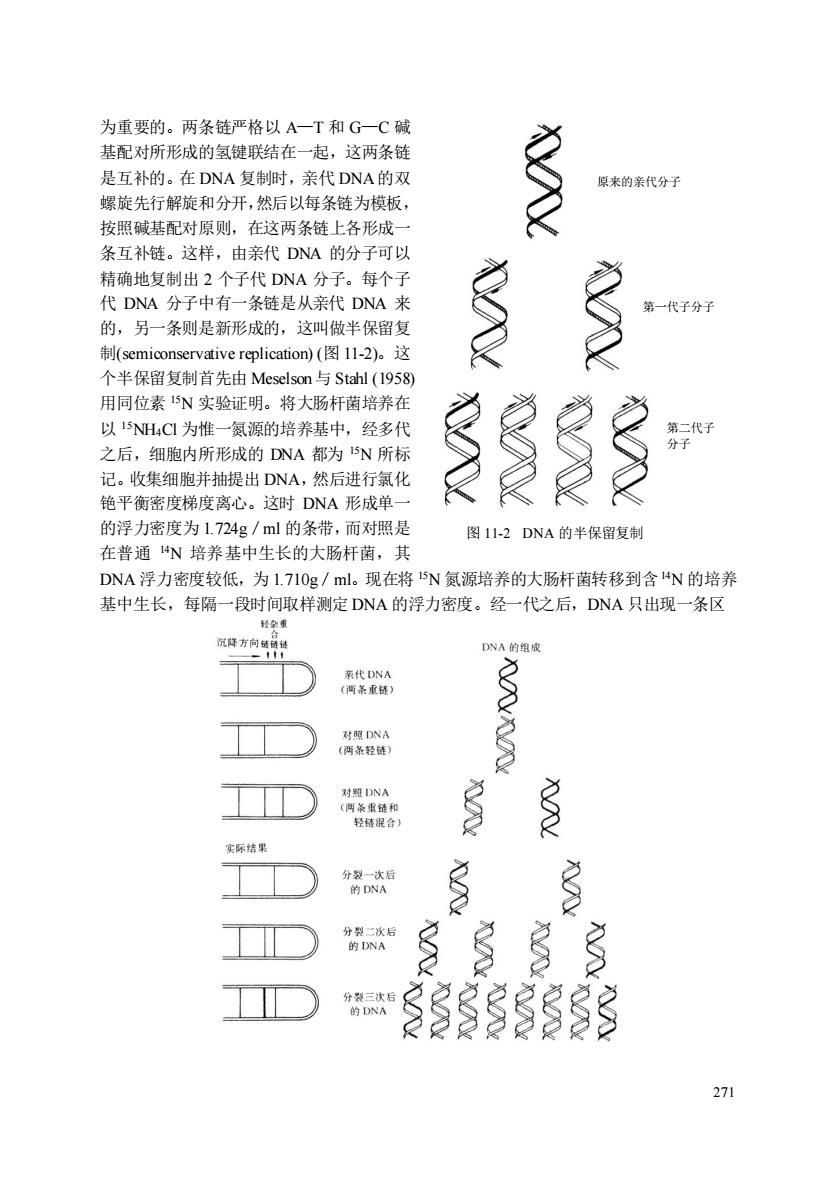
为重要的。两条链严格以A一T和G一C碱 基配对所形成的氢键联结在一起,这两条链 是互补的。在DNA复制时,亲代DNA的双 原来的亲代分 螺旋先行解旋和分开,然后以每条链为模板 按照碱基配对原则,在这两条链上各形城 条互补链。这样,由亲代DNA的分子可以 精确地复制出2个子代DNA分子。每个子 代DNA分子中有一条链是从亲代DNA来 的,另一条则是新形成的,这叫做半保留复 制(replication(图l-2).这 个半保留复制首先由Meselson与Stahl(1958) 用同位素5实验证明。将大肠杆菌培养在 以15NH4C1为惟一氮源的培养基中,经多代 之后,细胞内所形的DNA都为5N所材 记。收集细胞并抽提出DNA,然后进行氯化 铯平衡密度梯度离心。这时DNA形成单 的浮力密度为1.724g/ml的条带,而对照是 图1-2DNA的半保留复制 在普通N培养基中生长的大肠杆菌,其 DNA浮力密度较低,为1.71Og/ml。现在将15N氮源培养的大肠杆菌转移到含4N的培养 基中生长,每隔一段时间取样测定DNA的浮力密度。经一代之后, DNA只出现一条区 DNA的组 秀作 结果 分大 271
271 为重要的。两条链严格以 A—T 和 G—C 碱 基配对所形成的氢键联结在一起,这两条链 是互补的。在 DNA 复制时,亲代 DNA的双 螺旋先行解旋和分开,然后以每条链为模板, 按照碱基配对原则,在这两条链上各形成一 条互补链。这样,由亲代 DNA 的分子可以 精确地复制出 2 个子代 DNA 分子。每个子 代 DNA 分子中有一条链是从亲代 DNA 来 的,另一条则是新形成的,这叫做半保留复 制(semiconservative replication) (图 11-2)。这 个半保留复制首先由 Meselson与 Stahl (1958) 用同位素 15N 实验证明。将大肠杆菌培养在 以 15NH4Cl 为惟一氮源的培养基中,经多代 之后,细胞内所形成的 DNA 都为 15N 所标 记。收集细胞并抽提出 DNA,然后进行氯化 铯平衡密度梯度离心。这时 DNA 形成单一 的浮力密度为 1.724g/ml 的条带,而对照是 在普通 14N 培养基中生长的大肠杆菌,其 DNA 浮力密度较低,为 1.710g/ml。现在将 15N 氮源培养的大肠杆菌转移到含14N 的培养 基中生长,每隔一段时间取样测定 DNA 的浮力密度。经一代之后,DNA 只出现一条区 图 11-2 DNA 的半保留复制 原来的亲代分子 第二代子 分子 第一代子分子

图Il-3证明DNA半保留复制的Meselsen-Sahl实验图解 带,浮力密度为1.7I7g/ml,位于15N一DNA和14N一DNA之间,这条区带的DNA是由 4N/5N一DNA组成的。经二代之后,出现两条区带,其浮力密度分别为1.7I0g/ml和 1.717g/m,即一条区带为4N/I4N-DNA,另一条区带为14N/15N-DNA。再继续培养, 1N/1N一DNA分子逐渐增多,而1N/5N一DNA分子所占的比例逐渐减少。这些结果 及其解释可用图11-3表示。这个试验结果证明DNA是以半保留方式进行复制的。以后用 其他细菌、动物、植物、噬菌体、动物病毒等也证明了DNA的半保留复制。DNA的半保 留复制可以使遗传信息的传递保持相对的稳定,这和它的遗传功能是相吻合的,说明半保 留复制具有重要的生物学意义。但是这种稳定性是相对的。在一定条件下,DNA会发生损 伤,需要修复:在复制和转录中DN会有损耗,必须进行更新:在发育和分化过程中, DNA特定序列可能修饰、删除、扩增和重排 二、与DNA复制有关的酶和蛋白质 DNA的合成是以四种三磷酸脱氧核糖核苷为底物的聚合反应,该过程除了酶的催化 之外,还需要以适量的DNA为模板,以RNA或DNA)为引物和镁离子的参与。 mdATP、 DNA聚合酶 DNA+(m+m+m+na)PPi mdCTP DNA,Mg" ndTTP 实际上,DNA合成的反应是很复杂的,催化这个反应的酶也有多种,除DNA聚合酶 外,还有RNA引物合成酶(即引发酶),DNA连接酶、拓扑异构酶、解螺旋酶及多种蛋白 质因子参与。现将与DNA合成有关的几种酶和蛋白质因子扼要介绍如下: 1.引物合成酶(亦称引发酶,Primase) 此酶以DNA为模板合成一段RNA,这段RNA作为合成DNA的引物Primer)。催化 引物RNA合成的酶对利福平((rifampicin)不敏感,而且在一定程度上可用脱氧核糖核苷酸代 替核糖核苷酸作为底物,而与经典的RNA聚合酶不同。大肠杆菌的引物酶为一条单链多 肽,分子量为60000。 2.DNA聚合酶(DNA polymerase) 目前已知的DNA聚合酶有多种,它们的性状和在DNA合成中的功能均不相同。在大 肠杆菌中发现有3种DNA聚合酶,分别称为DNA聚合酶I、I、Ⅲ。DNA聚合酶I最初 是在1955年由Kornberg在大肠杆菌内发现的。Kornberg将其进行了高度纯化。纯化的酵 是一条单链多肽,呈球状,直径约为6.5m,是DNA直径的3倍左右。分子量为109000。 每个分子含一个锌原子。这个锌原子与酶的催化作用有关。DNA聚合酶I是多功能酶,它 具有5’→3'聚合酶,5’→3'外切酶及3′→5'外切酶的活性。它的主要功能是对 DNA损伤的修复,以及在DNA复制时,填补RNA引物切除后留下的空隙。 272
272 图 11-3 证明 DNA 半保留复制的 Meselsen-Stahl 实验图解 带,浮力密度为 1.7l7g/ml,位于 15N—DNA 和 14N—DNA 之间,这条区带的 DNA 是由 14N/15N—DNA 组成的。经二代之后,出现两条区带,其浮力密度分别为 1.710g/ml 和 1.717g/ml,即一条区带为 14N/14N—DNA,另一条区带为14N/15N -DNA。再继续培养, 14N/14N—DNA 分子逐渐增多,而 14N/15N—DNA 分子所占的比例逐渐减少。这些结果 及其解释可用图 11-3 表示。这个试验结果证明 DNA 是以半保留方式进行复制的。以后用 其他细菌、动物、植物、噬菌体、动物病毒等也证明了 DNA 的半保留复制。DNA 的半保 留复制可以使遗传信息的传递保持相对的稳定,这和它的遗传功能是相吻合的,说明半保 留复制具有重要的生物学意义。但是这种稳定性是相对的。在一定条件下,DNA 会发生损 伤,需要修复;在复制和转录中 DNA 会有损耗,必须进行更新;在发育和分化过程中, DNA 特定序列可能修饰、删除、扩增和重排。 二、与 DNA 复制有关的酶和蛋白质 DNA 的合成是以四种三磷酸脱氧核糖核苷为底物的聚合反应,该过程除了酶的催化 之外,还需要以适量的 DNA 为模板,以 RNA(或 DNA)为引物和镁离子的参与。 n1dATP + n2dGTP DNA 聚合酶 + DNA+ (n1+n2+n3+n4)PPi n3dCTP DNA, Mg2+ + n4dTTP 实际上,DNA合成的反应是很复杂的,催化这个反应的酶也有多种,除DNA 聚合酶 外,还有 RNA 引物合成酶(即引发酶),DNA 连接酶、拓扑异构酶、解螺旋酶及多种蛋白 质因子参与。现将与 DNA 合成有关的几种酶和蛋白质因子扼要介绍如下; 1. 引物合成酶(亦称引发酶,Primase) 此酶以 DNA 为模板合成一段 RNA,这段 RNA 作为合成 DNA 的引物(Primer)。催化 引物 RNA 合成的酶对利福平(rifampicin)不敏感,而且在一定程度上可用脱氧核糖核苷酸代 替核糖核苷酸作为底物,而与经典的 RNA 聚合酶不同。大肠杆菌的引物酶为一条单链多 肽,分子量为 60 000。 2. DNA 聚合酶(DNApolymerase) 目前已知的 DNA 聚合酶有多种,它们的性状和在 DNA 合成中的功能均不相同。在大 肠杆菌中发现有 3 种 DNA 聚合酶,分别称为 DNA 聚合酶 I、Ⅱ、Ⅲ。DNA 聚合酶I 最初 是在 1955 年由 Kornberg 在大肠杆菌内发现的。Kornberg 将其进行了高度纯化。纯化的酶 是一条单链多肽,呈球状,直径约为6.5nm,是 DNA 直径的 3 倍左右。分子量为 109 000。 每个分子含一个锌原子。这个锌原子与酶的催化作用有关。DNA 聚合酶 I 是多功能酶,它 具有 5′ →3′ 聚合酶,5′→3′外切酶及 3′→5′外切酶的活性。它的主要功能是对 DNA 损伤的修复,以及在 DNA 复制时,填补 RNA 引物切除后留下的空隙
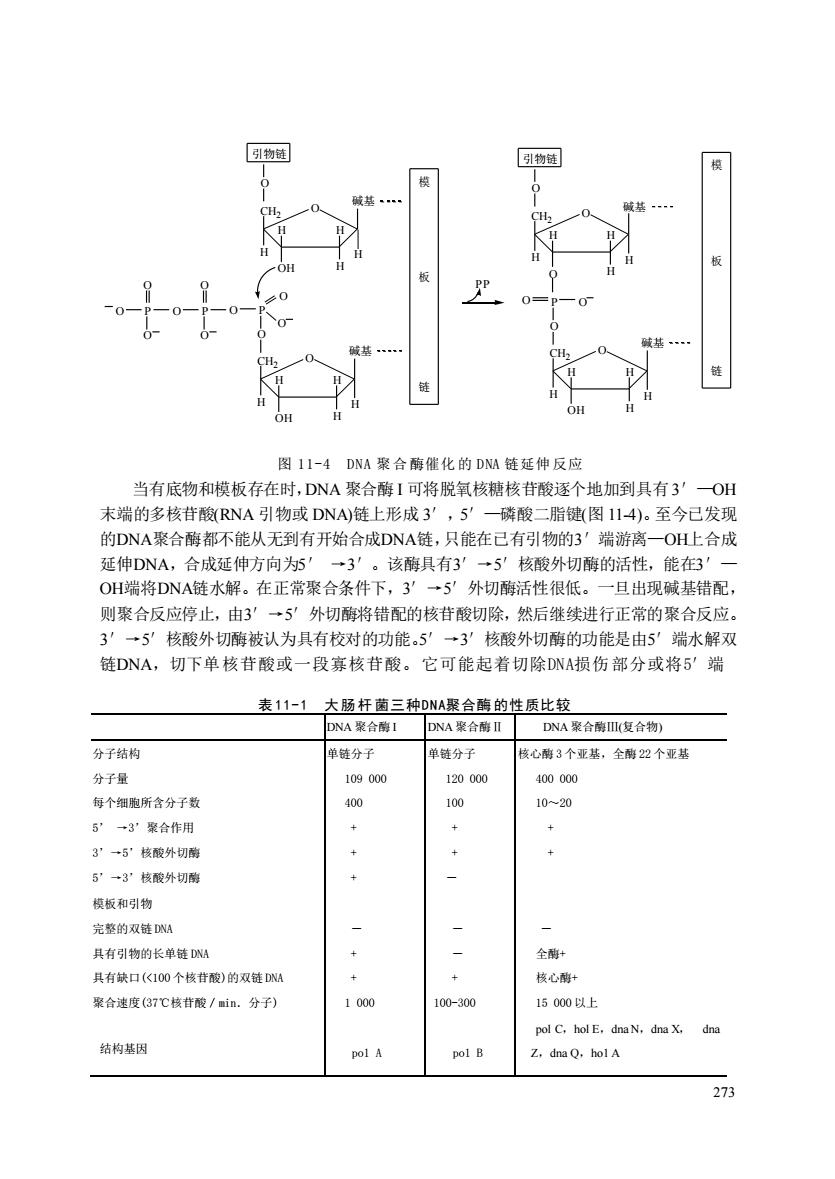
图II-4DNA聚合酶催化的DNM链延伸反应 当有底物和模板存在时,DNA聚合酶I可将脱氧核糖核苷酸逐个地加到具有3'OH 末端的多核苷酸RNA引物或DNA)链上形成3 ,5 一磷酸二脂键(图114)。至今已发现 的DNA聚合酶都不能从无到有开始合成DNA链,只能在己有引物的3'端游离一OH上合成 延伸DNA,合成延伸方向为5' →3'。该酶具有3一5'核酸外切酶的活性,能在3' OH端将DNA链水解。在正常聚合条件下,3'→5'外切酶活性很低。一旦出现碱基错配, 则聚合反应停止,由3'→5'外切酵将错配的核苷酸切除,然后继续进行正常的聚合反应。 3'→5'核酸外切酶被认为具有校对的功能。5'→3'核酸外切酶的功能是由5'端水解双 链DNA,切下单核苷酸或一段寡核苷酸。它可能起者切除DNA损伤部分或将5'端 表11-1 大肠杆菌三种DNA聚合酶的性质比较 DNA聚合醇I DNA案合酶I DNA聚合酶I(复合物) 分子结构 单链分子 单链分子 核心酶3个亚基,全酵2个亚基 分子量 109000 120000 400000 每个细胞所含分子数 1020 5”一3聚合作用 3”→5核酸外切酶 5”→3”核酸外切配 模板和引物 完整的双链DNA 其有引物的长单链D 全酶 具有缺口(《100个核作酸)的双链 + + 核心离+ 聚合速度(37℃核酸/in.分子) 1000 100-300 15000以上 pol C.holE,dnaN,dna X.dna 结构基因 pol A Z.dna Q.holA
273 图 11-4 DNA 聚 合 酶催化 的 DNA 链延伸 反应 当有底物和模板存在时,DNA 聚合酶 I 可将脱氧核糖核苷酸逐个地加到具有3′—OH 末端的多核苷酸(RNA 引物或 DNA)链上形成 3′,5′—磷酸二脂键(图 11-4)。至今已发现 的DNA聚合酶都不能从无到有开始合成DNA链,只能在已有引物的3′端游离—OH上合成 延伸DNA,合成延伸方向为5′ →3′。该酶具有3′→5′核酸外切酶的活性,能在3′— OH端将DNA链水解。在正常聚合条件下,3′→5′外切酶活性很低。一旦出现碱基错配, 则聚合反应停止,由3′→5′外切酶将错配的核苷酸切除,然后继续进行正常的聚合反应。 3′→5′核酸外切酶被认为具有校对的功能。5′→3′核酸外切酶的功能是由5′端水解双 链DNA,切下单核苷酸或一段寡核苷酸。它可能起着切除DNA损伤部分或将5′端 表 11-1 大 肠 杆 菌三种DNA聚合酶 的性质比较 DNA 聚合酶 I DNA 聚合酶Ⅱ DNA 聚合酶Ⅲ(复合物) 分子结构 分子量 每个细胞所含分子数 5’ →3’聚合作用 3’→5’核酸外切酶 5’→3’核酸外切酶 模板和引物 完整的双链 DNA 具有引物的长单链 DNA 具有缺口(<100 个核苷酸)的双链 DNA 聚合速度(37℃核苷酸/min.分子) 结构基因 单链分子 109 000 400 + + + - + + 1 000 po1 A 单链分子 120 000 100 + + - - - + 100-300 po1 B 核心酶 3 个亚基,全酶 22 个亚基 400 000 10~20 + + - 全酶+ 核心酶+ 15 000 以上 pol C,hol E,dna N,dna X, dna Z,dna Q,ho1 A CH2 H H OH H H H O CH2 H H OH H H H O O P O O O O P O O O P O O 碱基 碱基 O 引物链 模 板 链 CH2 H H O H H H O CH2 H H OH H H H O O O P O 碱基 碱基 O 引物链 模 板 链 PP
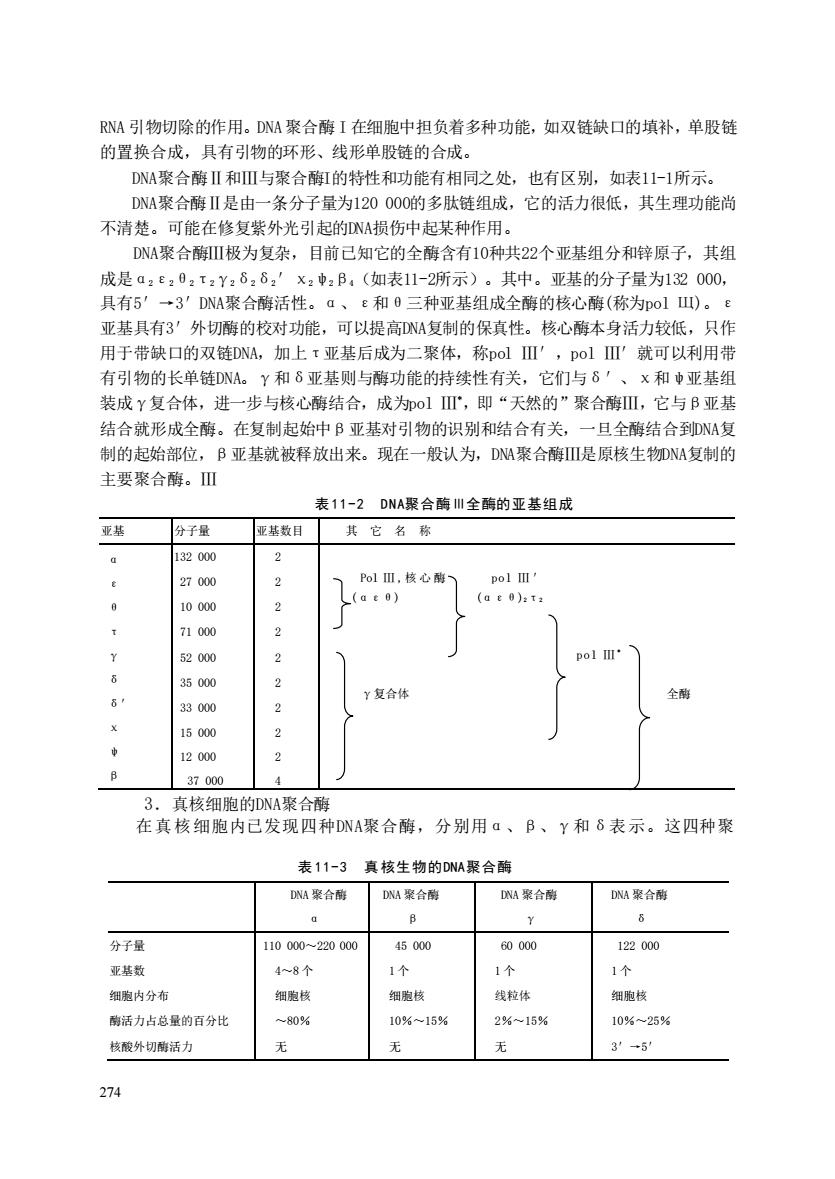
RNA引物切除的作用。DNA聚合酶I在细胞中担负者多种功能,如双链缺口的填补,单股链 的置换合成,具有引物的环形、线形单股链的合成。 DNA聚合酶Ⅱ和Ⅲ与聚合酶的特性和功能有相同之处,也有区别,如表11-1所示。 DA聚合酶Ⅱ是由一条分子量为120000的多肽链组成,它的活力很低,其生理功能尚 不清楚。可能在修复紫外光引起的DNA损伤中起某种作用。 DNA聚合酶Ⅲ极为复杂,目前己知它的全酶含有10种共22个亚基组分和锌原子,其组 成是a2e,0,t2Y2626,'x:中,B,(如表11-2所示)。其中。亚基的分子量为132000, 具有5'→3'DNA聚合酶活性。a、e和0三种亚基组成全酶的核心酶(称为PolI)。e 亚基具有3'外切酶的校对功能,可以提高NA复制的保真性。核心酶本身活力较低,只作 用于带缺口的双链DNA,加上τ亚基后成为二聚体,称polⅢ',pol'就可以利用带 有引物的长单链DNA。Y和δ亚基则与酶功能的特续性有关,它们与8'、 x和中亚基组 装成Y复合体,进一步与核心酶结合,成为o1Ⅲ',即“天然的”聚合酶Ⅲ,它与B亚基 结合就形成全酶。在复制起始中B亚基对引物的识别和结合有关,一旦全酶结合到DNA复 制的起始部位,B亚基就被释放出来。现在一般认为,DN州聚合酶是原核生物DNA复制的 主要聚合酶。Ⅲ 表11-2DNA聚合酶川全酶的亚基组成 亚基 分子量 亚基数目 其它名称 132000 27000 o1Ⅲ,核心酶、 10000 (at)t 71000 52000 pol II" 35000 Y复合体 全 3000 2 15000 2 12000 37000 3.真核细胞的DNA聚合酶 在真核细胞内己发现四种DNA聚合酶,分别用a、B、Y和6表示。这四种聚 表11-3真核生物的DNM聚合酶 DNA紧合梅 DNA聚合南 NA聚合酶 DNA紧合酶 分子量 110000220000 45000 60000 122000 亚基数 4~8个 1个 1个 细胞内分布 细胞核 细胞核 线拉体 细胞核 酶活力占总量的百分比 -80% 10%~15% 2%~15% 10%~25% 核酸外切酶活力 无 无 3'→5 274
274 RNA 引物切除的作用。DNA聚合酶 I 在细胞中担负着多种功能,如双链缺口的填补,单股链 的置换合成,具有引物的环形、线形单股链的合成。 DNA聚合酶Ⅱ和Ⅲ与聚合酶I的特性和功能有相同之处,也有区别,如表11-1所示。 DNA聚合酶Ⅱ是由一条分子量为120 000的多肽链组成,它的活力很低,其生理功能尚 不清楚。可能在修复紫外光引起的DNA损伤中起某种作用。 DNA聚合酶Ⅲ极为复杂,目前已知它的全酶含有10种共22个亚基组分和锌原子,其组 成是α2ε2θ2τ2γ2δ2δ2′χ2ψ2β4(如表11-2所示)。其中。亚基的分子量为132 000, 具有5′→3′DNA聚合酶活性。α、ε和θ三种亚基组成全酶的核心酶(称为pol Щ)。ε 亚基具有3′外切酶的校对功能,可以提高DNA复制的保真性。核心酶本身活力较低,只作 用于带缺口的双链DNA,加上τ亚基后成为二聚体,称pol Ⅲ′,pol Ⅲ′就可以利用带 有引物的长单链DNA。γ和δ亚基则与酶功能的持续性有关,它们与δ′、χ和ψ亚基组 装成γ复合体,进一步与核心酶结合,成为pol Ⅲ *,即“天然的”聚合酶Ⅲ,它与β亚基 结合就形成全酶。在复制起始中β亚基对引物的识别和结合有关,一旦全酶结合到DNA复 制的起始部位,β亚基就被释放出来。现在一般认为,DNA聚合酶Ⅲ是原核生物DNA复制的 主要聚合酶。Ⅲ 表 11-2 DNA聚合酶Ⅲ全酶的亚基组成 3.真核细胞的DNA聚合酶 在真核细胞内已发现四种DNA聚合酶,分别用α、β、γ和δ表示。这四种聚 表 11-3 真 核生物的DNA聚合酶 亚基 分子量 亚基数目 其 它 名 称 α ε θ τ γ δ δ′ χ ψ β 132 000 27 000 10 000 71 000 52 000 35 000 33 000 15 000 12 000 37 000 2 2 2 2 2 2 2 2 2 4 Pol Ⅲ ,核心酶 pol Ⅲ ′ (αεθ) (αεθ)2τ2 pol Ⅲ * γ复合体 全酶 DNA 聚合酶 α DNA 聚合酶 β DNA 聚合酶 γ DNA 聚合酶 δ 分子量 亚基数 细胞内分布 酶活力占总量的百分比 核酸外切酶活力 110 000~220 000 4~8 个 细胞核 ~80% 无 45 000 1 个 细胞核 10%~15% 无 60 000 1 个 线粒体 2%~15% 无 122 000 1 个 细胞核 10%~25% 3′→5′