
合酶的特性见表11-3。现在一般认为DNA聚合酶a和6的作用是复制染色体DNA,主要的根 据是它们在细胞内活力水平的变化与DA复制有明显的平行关系,在分裂细胞的S期达到高 峰。聚合酶ā催化随后链的合成,而聚合酶6催化领头链的合成,它还具有3'→5'外切 酶的活力。NA聚合酶B的功能主要是修复作用。NA聚合爵Y是从线粒体中分离得到的, 推测它与线粒体DNA的复制有关。 4.DNA连接酶(DNM1 igase) 其作用是催化双链DNA中的切口处的相邻5'一磷酸基与3'一羟基之间形成磷酸酯键。 但是它不能将两条游离的DNA单链连接起来 大肠杆菌的DNA连接酶要求NAD提供能量,产物是AMP和烟酰胺单核苷酸(图11-5)。而在 高等生物中,则要求ATP提供能量,产物是AMP和焦磷酸。大肠杆菌的DNA连接酶是分子量为 75000的多肽链。在哺乳动物细胞中发现至少有两种连接酵,分别称为连接酶I和Ⅱ。连接 酶I的分子量为200000,连接酶Ⅱ的为85000。连接酶主要在正在繁殖的细胞中起作用。 连接酶Ⅱ则在停止分裂的细胞中起作用。DNA连接酶在DNA复制、修复、重组中均起重要作 用。 E+ATR或NAD EAMP+PP或NMN E-AMP+-5-DNA= E+AMP-(B-5'-DNA DNA-3-OH+AMP-5-DNA= =DNA-3-0-5'-DNA+AMP DNA-3OH+®-5DNA+ATP或NAD) =DNA-3O-⊙-S-DNA+AMP+PP或NMN 图11-5DNA连接酶催化反应的机理 5.拓扑异构酶(topoisomerase) 生物体内DNA分子通常处于超螺旋状态,而DNA的许多生物功能需要解开双链才能进 行。拓扑异构酶就是催化DNA的拓扑连环数发生变化的酶,它可分为拓扑异构酶和拓扑 异构酶Ⅱ(也称为旋转酶,gyrase)。1型酶可使双链DNA分子中的一条链发生断裂和再连接, 反应不需要提供能量,它们主要集中在活性转录区,同转录有关。Ⅱ型酶能使DN八的两条 链同时发生断裂和再连接,当它引入负超螺旋时需要由ATP提供能量。一个拓扑异构酶Ⅱ 的分子1mi分钟可引入100个负超螺旋。它们主要分布在染色质骨架蛋白和核基质部位, 同复制有关。拓扑异构酶可减少负超螺旋:拓扑异构酶红可引入负超螺旋,它们协同作用 控制着DNA的拓扑结构。拓扑异构酶在重组、修复和DNA的其他转变方面起着重要的作用。 6.解螺旋與helicase) 这类酶能通过水解ATP将DNA的两条链打开。ATP水解活力要有单链DNA存在时才表 现。大肠杆菌中的rep蛋白(rep基因的产物就是这样一种酶。由分子量为65000的一条多肽 链组成。每解开一对碱基需要水解2个ATP分子。 7.其他蛋白因子 (I)单链结合蛋白SSP)它的功能是稳定已被解开的DNA单链,阻止复性和保护单 链不被核酸酶降解。 (2)引发前体(preprimosome)它是由6种蛋白质即dnaB、dhaC、n、n'、n”和i组 成。引发前体再与RNA引物合成酶(引发酶)结合,组装成引发体(primosome)。引发体结 275
275 合酶的特性见表11-3。现在一般认为DNA聚合酶α和δ的作用是复制染色体DNA,主要的根 据是它们在细胞内活力水平的变化与DNA复制有明显的平行关系,在分裂细胞的S期达到高 峰。聚合酶α催化随后链的合成,而聚合酶δ催化领头链的合成,它还具有3′→5′外切 酶的活力。DNA聚合酶β的功能主要是修复作用。DNA聚合酶γ是从线粒体中分离得到的, 推测它与线粒体DNA的复制有关。 4.DNA连接酶(DNA ligase) 其作用是催化双链DNA中的切口处的相邻5′—磷酸基与3′—羟基之间形成磷酸酯键。 但是它不能将两条游离的DNA单链连接起来。 大肠杆菌的DNA连接酶要求NAD+提供能量,产物是AMP和烟酰胺单核苷酸(图11-5)。而在 高等生物中,则要求ATP提供能量,产物是AMP和焦磷酸。大肠杆菌的DNA连接酶是分子量为 75 000的多肽链。在哺乳动物细胞中发现至少有两种连接酶,分别称为连接酶I和Ⅱ。连接 酶I的分子量为200 000,连接酶Ⅱ的为85 000。连接酶I主要在正在繁殖的细胞中起作用。 连接酶Ⅱ则在停止分裂的细胞中起作用。DNA连接酶在DNA复制、修复、重组中均起重要作 用。 图 11-5 DNA连接酶催化反应的机理 5.拓扑异构酶(topoisomerase) 生物体内DNA分子通常处于超螺旋状态,而DNA的许多生物功能需要解开双链才能进 行。拓扑异构酶就是催化DNA的拓扑连环数发生变化的酶,它可分为拓扑异构酶I和拓扑 异构酶Ⅱ(也称为旋转酶,gyrase)。I型酶可使双链DNA分子中的一条链发生断裂和再连接, 反应不需要提供能量,它们主要集中在活性转录区,同转录有关。Ⅱ型酶能使DNA的两条 链同时发生断裂和再连接,当它引入负超螺旋时需要由ATP提供能量。一个拓扑异构酶Ⅱ 的分子1min分钟可引入100个负超螺旋。它们主要分布在染色质骨架蛋白和核基质部位, 同复制有关。拓扑异构酶I可减少负超螺旋;拓扑异构酶II可引入负超螺旋,它们协同作用 控制着DNA的拓扑结构。拓扑异构酶在重组、修复和DNA的其他转变方面起着重要的作用。 6.解螺旋酶(helicase) 这类酶能通过水解ATP将DNA的两条链打开。ATP水解活力要有单链DNA存在时才表 现。大肠杆菌中的rep蛋白(rep基因的产物)就是这样一种酶。由分子量为65 000的一条多肽 链组成。每解开一对碱基需要水解2个ATP分子。 7.其他蛋白因子 (1)单链结合蛋白(SSP) 它的功能是稳定已被解开的DNA单链,阻止复性和保护单 链不被核酸酶降解。 (2)引发前体(preprimosome) 它是由6种蛋白质即dnaB、dnaC、n、n′、n″和i组 成。引发前体再与RNA引物合成酶(引发酶)结合,组装成引发体(primosome)。引发体结 NAD +) P P P P + AMP P NAD +) P +AMP + PPi(或NMN) E + AT(P 或 E·AMP + PPi(或NMN) E-AMP + 5'-DNA E + AMP 5'-DNA DNA-3'-OH + AMP 5'-DNA DNA-3'-O 5'-DNA DNA-3'-OH + 5'-DNA + AT(P 或 DNA-3'-O 5'-DNA
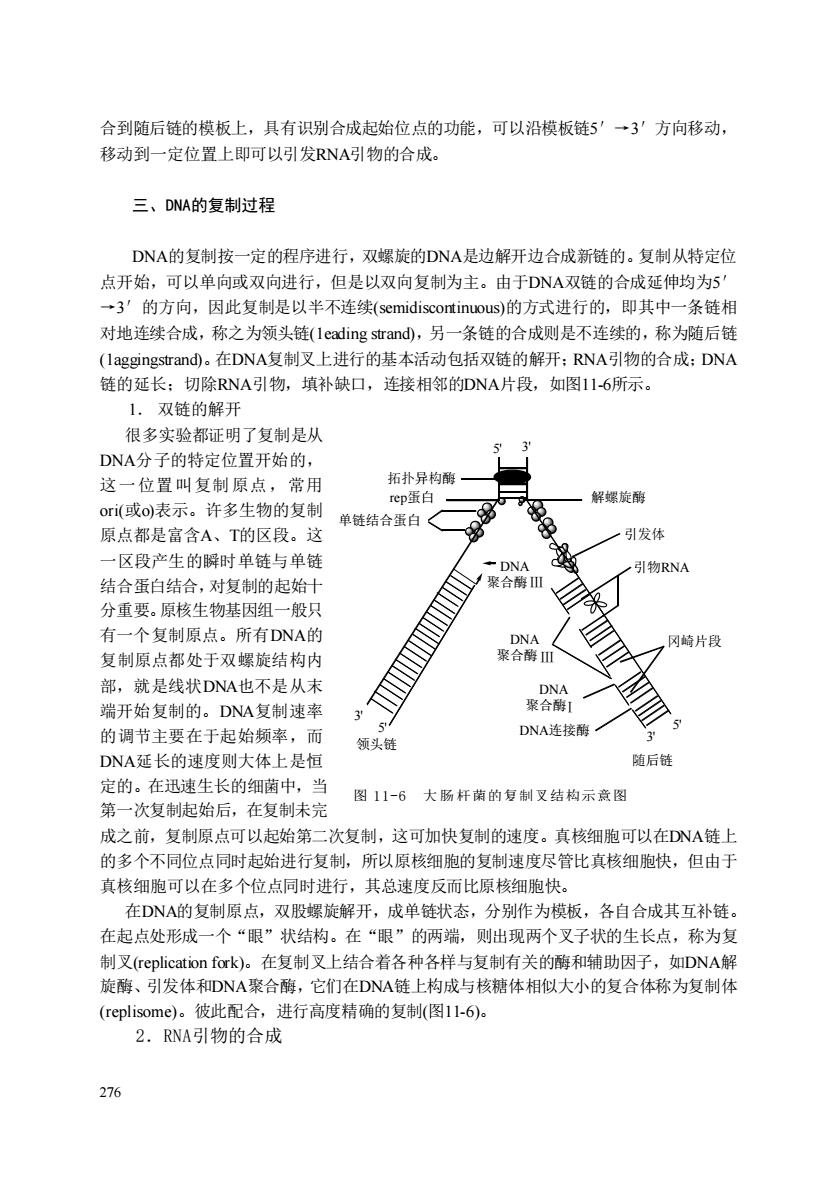
合到随后链的模板上,具有识别合成起始位点的功能,可以沿模板链'→3'方向移动, 移动到一定位置上即可以引发RNA引物的合成。 三、DNA的复制过程 DNA的复制按一定的程序进行,双螺旋的DNA是边解开边合成新链的。复制从特定位 点开始,可以单向或双向进行,但是以双向复制为主。由于DNA双链的合成延伸均为5 一3'的方向,因此复制是以半不连续))的方式进行的,即其中一条链相 对地连续合成,称之为领头链(leading strand),另一条链的合成则是不连续的,称为随后链 (laggingstrand)。在DNA复制叉上进行的基本活动包括双链的解开:RNA引物的合成:DNA 的延长:切除RNA引物,填补缺口,连接相邻的DNA片段,如图11-6所示。 1.双链的解开 很多实验都证明了复制是从 DNA分子的特定位置开始的, 这一位署叫复制原点,常用 拓扑异构酶 ori(或o)表示。许多生物的复制 解螺刷 单链结合蛋白 原点都是富含A、T的区段。这 引发体 一区段产生的瞬时单链与单链 引物RNA 结合蛋白结合,对复制的起始十 √聚合酸川 分重要。原核生物基因组一般只 有一个复制原点。所有DNA的 聚m ,冈崎片段 复制原点都处于双螺旋结构内 部,就是线状DNA也不是从未 端开始复制的。DNA复制速率 3 的调节主要在于起始频率, 而 DNA连接南 领头特 DNA延长的速度则大体上是恒 随后链 定的。在讯谏生长的细菌中,当 图11-6大肠杆菌的复制叉结构示意图 第一次复制起始后,在复制未完 成之前.复制原占可以起始第二次复制.这可加快复制的速府。直核细胞口可以在D八NA钵 的多个不同位点同时起始进行复制,所以原核细胞的复制速度尽管比真核细胞快,但由于 直核细朐响可以在名个位占同时讲行,其总谏度反而比原核细朐快 在DNA的复制原点,双股螺旋解开,成单链状态,分别作为模板,各自合成其互补链。 在起点处形成一个“眼”状结构。在“眼”的两端,则出现两个叉子状的生长点,称为复 制叉(replication fork)。在复制叉上结合着各种各样与复制有关的酶和辅助因子,如DNA解 旋酶、引发体和DNA聚合酶,它们在DNA链上构成与核糖体相似大小的复合体称为复制体 (replisome)。彼此配合,进行高度精确的复制(图1l-6)。 2.RNA引物的合成 276
276 合到随后链的模板上,具有识别合成起始位点的功能,可以沿模板链5′→3′方向移动, 移动到一定位置上即可以引发RNA引物的合成。 三、DNA的复制过程 DNA的复制按一定的程序进行,双螺旋的DNA是边解开边合成新链的。复制从特定位 点开始,可以单向或双向进行,但是以双向复制为主。由于DNA双链的合成延伸均为5′ →3′的方向,因此复制是以半不连续(semidiscontinuous)的方式进行的,即其中一条链相 对地连续合成,称之为领头链(1eading strand),另一条链的合成则是不连续的,称为随后链 (1aggingstrand)。在DNA复制叉上进行的基本活动包括双链的解开;RNA引物的合成;DNA 链的延长;切除RNA引物,填补缺口,连接相邻的DNA片段,如图11-6所示。 1. 双链的解开 很多实验都证明了复制是从 DNA分子的特定位置开始的, 这一位置叫复制原点,常用 ori(或o)表示。许多生物的复制 原点都是富含A、T的区段。这 一区段产生的瞬时单链与单链 结合蛋白结合,对复制的起始十 分重要。原核生物基因组一般只 有一个复制原点。所有DNA的 复制原点都处于双螺旋结构内 部,就是线状DNA也不是从末 端开始复制的。DNA复制速率 的调节主要在于起始频率,而 DNA延长的速度则大体上是恒 定的。在迅速生长的细菌中,当 第一次复制起始后,在复制未完 成之前,复制原点可以起始第二次复制,这可加快复制的速度。真核细胞可以在DNA链上 的多个不同位点同时起始进行复制,所以原核细胞的复制速度尽管比真核细胞快,但由于 真核细胞可以在多个位点同时进行,其总速度反而比原核细胞快。 在DNA的复制原点,双股螺旋解开,成单链状态,分别作为模板,各自合成其互补链。 在起点处形成一个“眼”状结构。在“眼”的两端,则出现两个叉子状的生长点,称为复 制叉(replication fork)。在复制叉上结合着各种各样与复制有关的酶和辅助因子,如DNA解 旋酶、引发体和DNA聚合酶,它们在DNA链上构成与核糖体相似大小的复合体称为复制体 (replisome)。彼此配合,进行高度精确的复制(图11-6)。 2.RNA引物的合成 图 11-6 大 肠 杆菌的复制叉结构示意图 5' 3' 5' 3' 5' 3' 拓扑异构酶 rep蛋白 单链结合蛋白 解螺旋酶 引发体 冈崎片段 Ⅰ Ⅲ DNA 聚合酶 Ⅲ DNA 聚合酶 DNA 聚合酶 DNA连接酶 领头链 随后链 引物RNA

在DA复制的起始处双链解开,领头链先引发开始合成,与其模板形成双链结构,而另 条亲代链则被置换出来。只有在领头链将另一条亲本链的特别序列置换出来,才能产生 随后链的前体片段的前引发作用。需要引发酶与引发前体结合形成引发体。引发体在复制 叉上移动,识别合成的起始点,引发RNA引物的合成。移动和引发均需要由ATP提供能量, 以DNA为模板,按5'一3'的方向,合成一段引物RNA链。引物长度约为几个至10个核苷酸。 在引物的5'端含3个磷酸残基,3'端为游离的羟基。 3.DNA链的延长 当RNA引物合成之后,在DNA聚合酶的催化下,以四种脱氧核糖核苷5'-三磷酸为底 物,在RNA引物的3'端以磷酸二酯键连接上脱氧核糖核苷酸并释放出PPi。DNA链的合成是 以两条亲代DNA链为模板,按碱基配对原则进行复制的,亲代NA的双股链呈反向平行, 条链是5'◆3'方向,另一条链是3”◆5方向。在个复制叉内两条链的复制方向不同 (图11-7),所以新合成的二条子链极性也正好相反。由于迄今为止还没有发现一种DN聚合 酶能按3'→5'方向延伸,因此子链中有一条链沿着亲代DNA单链的3'→5'方向(亦即 新合成的DNA沿5'→3'方向)不断延长。这条新链称为领头链。而另一条链的合成方向 与复制叉的前进方向相反,只能断续地合成5'→3'的多个短片段。1968年冈崎发现了这 些片段故又称为冈崎片段(Okazaki fragment)。它们随后连接成大片段,这条新链称为随后 链。这种领头链是连续合成的,随后链断续合成的方式称为半不连续复制(semidiscontinuous replication。)原核细胞的冈崎片段长度为1000~2000个核苷酸。真核细胞的较短,长度为 100200个核苷酸。 复制 Lm 随后链 儿领头鼓 氧头链 随后链 延伸 WΠΠ 延伸 起点 图II-7DNA的双向复制 尽管领头链的合成总是领先一段,但是从来没有发现领头链跑得太远,而总是与随后 链保持相对稳定的一段距离。988年Kornberg等人从大肠杆菌中分离出8O0kDa的polⅢ 和900kDa的polⅢ全酶,其中各个亚基均有两个,并证明是具有双活性部位的非对称结构, 这表明同一个p©Ⅲ全酶可能同时负责领头链和随后链的复制。根据实验资料提出如下复 制模型(图11-8),随后链的模板在DNA聚合酶Ⅲ全酶上绕转180°形城一个小环,使冈崎片 段的合成方向能够与领头链的合成方向以及复制体的移动方向保持一致。随着冈崎片段的 延长,这个大环从DNA聚合酶上释放。在此之前,领头链的合成已将另一部分随后链的 模板置换出来,并在适于引发的位点上由引发体合成新的引物,然后再形成一个小环而进 行新的冈崎片段的合成。 277
277 在DNA复制的起始处双链解开,领头链先引发开始合成,与其模板形成双链结构,而另 一条亲代链则被置换出来。只有在领头链将另一条亲本链的特别序列置换出来,才能产生 随后链的前体片段的前引发作用。需要引发酶与引发前体结合形成引发体。引发体在复制 叉上移动,识别合成的起始点,引发RNA引物的合成。移动和引发均需要由ATP提供能量。 以DNA为模板,按5′→3′的方向,合成一段引物RNA链。引物长度约为几个至10个核苷酸。 在引物的5′端含3个磷酸残基,3′端为游离的羟基。 3.DNA链的延长 当RNA引物合成之后,在DNA聚合酶Ⅲ的催化下,以四种脱氧核糖核苷5′-三磷酸为底 物,在RNA引物的3′端以磷酸二酯键连接上脱氧核糖核苷酸并释放出PPi。DNA链的合成是 以两条亲代DNA链为模板,按碱基配对原则进行复制的,亲代DNA的双股链呈反向平行,一 条链是5′→3′方向,另一条链是3′→5′方向。在一个复制叉内两条链的复制方向不同 (图11-7),所以新合成的二条子链极性也正好相反。由于迄今为止还没有发现一种DNA聚合 酶能按3′→5′方向延伸,因此子链中有一条链沿着亲代DNA单链的3′→5′方向(亦即 新合成的DNA沿5′→3′方向)不断延长。这条新链称为领头链。而另一条链的合成方向 与复制叉的前进方向相反,只能断续地合成5′→3′的多个短片段。1968年冈崎发现了这 些片段故又称为冈崎片段(Okazaki fragment)。它们随后连接成大片段,这条新链称为随后 链。这种领头链是连续合成的,随后链断续合成的方式称为半不连续复制(semidiscontinuous replication)。原核细胞的冈崎片段长度为1 000~2 000个核苷酸。真核细胞的较短,长度为 100~200个核苷酸。 随后链 领头链 复制叉 延伸 3' 5' 起点 随后链 领头链 复制叉 延伸 3' 5' 起点 3' 3' 3' 3' 图11-7 DNA的双向复制 尽管领头链的合成总是领先一段,但是从来没有发现领头链跑得太远,而总是与随后 链保持相对稳定的一段距离。1988年Kornberg等人从大肠杆菌中分离出800 kDa的pol Ⅲ* 和900 kDa的pol Ⅲ全酶,其中各个亚基均有两个,并证明是具有双活性部位的非对称结构, 这表明同一个pol Ⅲ全酶可能同时负责领头链和随后链的复制。根据实验资料提出如下复 制模型(图11-8),随后链的模板在DNA聚合酶Ⅲ全酶上绕转180º形成一个小环,使冈崎片 段的合成方向能够与领头链的合成方向以及复制体的移动方向保持一致。随着冈崎片段的 延长,这个大环从DNA聚合酶Ⅲ上释放。在此之前,领头链的合成已将另一部分随后链的 模板置换出来,并在适于引发的位点上由引发体合成新的引物,然后再形成一个小环而进 行新的冈崎片段的合成