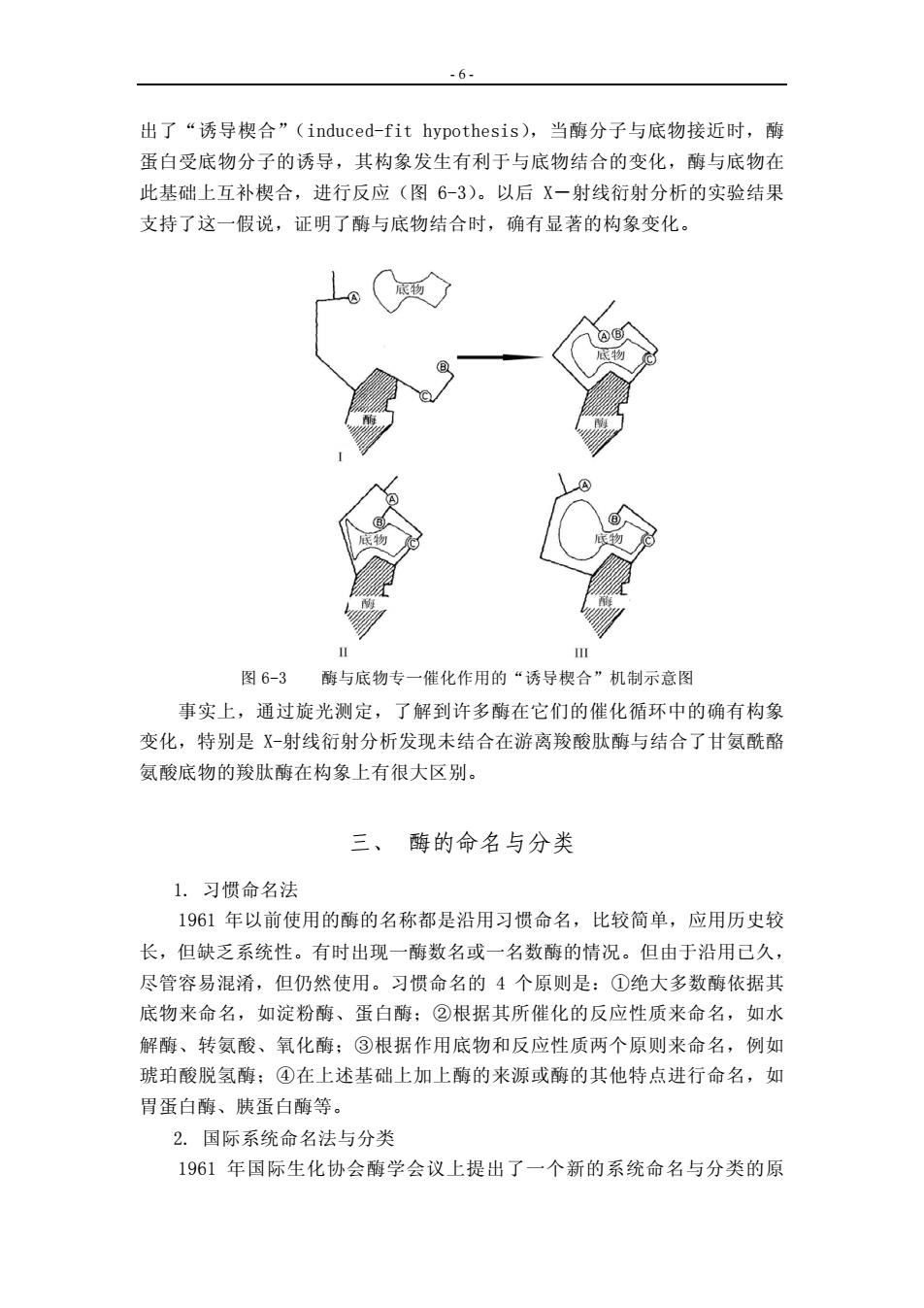
- 6 - 出了“诱导楔合”(induced-fit hypothesis),当酶分子与底物接近时,酶 蛋白受底物分子的诱导,其构象发生有利于与底物结合的变化,酶与底物在 此基础上互补楔合,进行反应(图 6-3)。以后 X-射线衍射分析的实验结果 支持了这一假说,证明了酶与底物结合时,确有显著的构象变化。 图 6-3 酶与底物专一催化作用的“诱导楔合”机制示意图 事实上,通过旋光测定,了解到许多酶在它们的催化循环中的确有构象 变化,特别是 X-射线衍射分析发现未结合在游离羧酸肽酶与结合了甘氨酰酪 氨酸底物的羧肽酶在构象上有很大区别。 三、 酶的命名与分类 1. 习惯命名法 1961 年以前使用的酶的名称都是沿用习惯命名,比较简单,应用历史较 长,但缺乏系统性。有时出现一酶数名或一名数酶的情况。但由于沿用已久, 尽管容易混淆,但仍然使用。习惯命名的 4 个原则是:①绝大多数酶依据其 底物来命名,如淀粉酶、蛋白酶;②根据其所催化的反应性质来命名,如水 解酶、转氨酸、氧化酶;③根据作用底物和反应性质两个原则来命名,例如 琥珀酸脱氢酶;④在上述基础上加上酶的来源或酶的其他特点进行命名,如 胃蛋白酶、胰蛋白酶等。 2. 国际系统命名法与分类 1961 年国际生化协会酶学会议上提出了一个新的系统命名与分类的原
- 6 - 出了“诱导楔合”(induced-fit hypothesis),当酶分子与底物接近时,酶 蛋白受底物分子的诱导,其构象发生有利于与底物结合的变化,酶与底物在 此基础上互补楔合,进行反应(图 6-3)。以后 X-射线衍射分析的实验结果 支持了这一假说,证明了酶与底物结合时,确有显著的构象变化。 图 6-3 酶与底物专一催化作用的“诱导楔合”机制示意图 事实上,通过旋光测定,了解到许多酶在它们的催化循环中的确有构象 变化,特别是 X-射线衍射分析发现未结合在游离羧酸肽酶与结合了甘氨酰酪 氨酸底物的羧肽酶在构象上有很大区别。 三、 酶的命名与分类 1. 习惯命名法 1961 年以前使用的酶的名称都是沿用习惯命名,比较简单,应用历史较 长,但缺乏系统性。有时出现一酶数名或一名数酶的情况。但由于沿用已久, 尽管容易混淆,但仍然使用。习惯命名的 4 个原则是:①绝大多数酶依据其 底物来命名,如淀粉酶、蛋白酶;②根据其所催化的反应性质来命名,如水 解酶、转氨酸、氧化酶;③根据作用底物和反应性质两个原则来命名,例如 琥珀酸脱氢酶;④在上述基础上加上酶的来源或酶的其他特点进行命名,如 胃蛋白酶、胰蛋白酶等。 2. 国际系统命名法与分类 1961 年国际生化协会酶学会议上提出了一个新的系统命名与分类的原

- 7 - 则,已为国际生化协会所采用。1992 年对此进行了修改。按照国际系统命名 法原则,每一种酶有一个系统名称和习惯名称,前者必须明确标准酶的底物 及催化反应的性质,每个酶有一个特定的编号。这种系统命名原则及系统编 号是相当严格的。 国际系统分类的原则是将所有的酶促反应按反应性质分为 6 大类,分别 用 1、2、3、4、5、6 的编号表示,再根据底物中被作用的基团或键的特点将 每一大类分为若干个亚类,每一个亚类又按顺序编成 1、2、3、4.等数字, 每一个亚类可再分为若干个亚-亚类,仍用 1、2、3、4.编号。每一种酶都 被指定为一个由 4 位数字组成的酶委员会编号(EC number)。例如黄嘌呤氧 化还原酶的系统命名为 EC 1.2.3.2。其中第一个数字指明该酶属于 6 大类中 的哪一类;第二个数为酶属的亚类;第三个数字是该酶所属的亚-亚类;第 四个数字则表明该酶在一定亚-亚类中的排号。-表 6-2 列出了食品中的一 些重要酶的系统分类。 表 6-2 食品中的一些重要的酶分类 类和亚类 酶 EC 编号 1. 氧化还原酶 1.1 供体为 OH-OH 1.1.1 受体为NAD+ 或NADP+ 乙醇脱氢酶 1.1.1.1 丁二醇脱氢酶 1.1.1.4 L-艾杜糖醇-2-脱氢酶 1.1.1.14 L-乳糖脱氢酶 1.1.1.27 苹果酸脱氢酶 1.1.1.37 半乳糖-1-脱氢酶 1.1.1.48 葡萄糖-6-磷酸-1-脱氢酶 1.1.1.49 1.1.3 受体为氧 葡萄糖氧化酶 1.1.3.4 黄嘌呤氧化酶 1.1.3.22 1.2 供体为醛基 1.2.1 受体为NAD+ 或NADP+ 醛脱氢酶 1.2.1.3 1.8 供体为含硫化物 1.8.5 受体为醌或醌类化合物 谷胱甘肽脱氢酶 1.8.5.1 1.10 体供为二烯醇或二酚 1.10.3 受体为氧 抗坏血酸氧化酶 1.10.3.3 1.11 受体为氢过化物 过氧化氢酶 1.11.1.6 过氧化物酶 1.11.1.7 1.13 作用于单一供体 1.13.11 与分子氧结合 脂肪氧合酶 1.13.11.12
- 7 - 则,已为国际生化协会所采用。1992 年对此进行了修改。按照国际系统命名 法原则,每一种酶有一个系统名称和习惯名称,前者必须明确标准酶的底物 及催化反应的性质,每个酶有一个特定的编号。这种系统命名原则及系统编 号是相当严格的。 国际系统分类的原则是将所有的酶促反应按反应性质分为 6 大类,分别 用 1、2、3、4、5、6 的编号表示,再根据底物中被作用的基团或键的特点将 每一大类分为若干个亚类,每一个亚类又按顺序编成 1、2、3、4.等数字, 每一个亚类可再分为若干个亚-亚类,仍用 1、2、3、4.编号。每一种酶都 被指定为一个由 4 位数字组成的酶委员会编号(EC number)。例如黄嘌呤氧 化还原酶的系统命名为 EC 1.2.3.2。其中第一个数字指明该酶属于 6 大类中 的哪一类;第二个数为酶属的亚类;第三个数字是该酶所属的亚-亚类;第 四个数字则表明该酶在一定亚-亚类中的排号。-表 6-2 列出了食品中的一 些重要酶的系统分类。 表 6-2 食品中的一些重要的酶分类 类和亚类 酶 EC 编号 1. 氧化还原酶 1.1 供体为 OH-OH 1.1.1 受体为NAD+ 或NADP+ 乙醇脱氢酶 1.1.1.1 丁二醇脱氢酶 1.1.1.4 L-艾杜糖醇-2-脱氢酶 1.1.1.14 L-乳糖脱氢酶 1.1.1.27 苹果酸脱氢酶 1.1.1.37 半乳糖-1-脱氢酶 1.1.1.48 葡萄糖-6-磷酸-1-脱氢酶 1.1.1.49 1.1.3 受体为氧 葡萄糖氧化酶 1.1.3.4 黄嘌呤氧化酶 1.1.3.22 1.2 供体为醛基 1.2.1 受体为NAD+ 或NADP+ 醛脱氢酶 1.2.1.3 1.8 供体为含硫化物 1.8.5 受体为醌或醌类化合物 谷胱甘肽脱氢酶 1.8.5.1 1.10 体供为二烯醇或二酚 1.10.3 受体为氧 抗坏血酸氧化酶 1.10.3.3 1.11 受体为氢过化物 过氧化氢酶 1.11.1.6 过氧化物酶 1.11.1.7 1.13 作用于单一供体 1.13.11 与分子氧结合 脂肪氧合酶 1.13.11.12

- 8 - 1.14 作用于一对供体 一元酚单加氧酶(多酚氧化酶) 1.14.18.1 2 转移酶 2.7 转移磷酸 2.7.1 受体为 OH 己糖激酶 2.7.1.1 甘油激酶 2.7.1.30 丙酮酸激酶 2.7.1.40 2.7.3 受体为 N-基 肌酸激酶 2.7.3.2 3 水解酶 3.1 切断酯键 3.1.1 羧酸酯水解酶 羧酸酯酶 3.1.1.1 三酰甘油酯酶 3.1.1.3 磷酸酯酶A2 3.1.1.4 乙酰胆碱酯酶 3.1.1.7 果胶甲酯酶 3.1.1.11 磷酸酯酶A1 3.1.1.32 3.1.3 磷酸单酯水解酶 碱性磷酸酯酶 3.1.3.1 3.1.4 磷酸双酯水解酶 磷脂酶 C 3.1.4.2 磷脂酶 D 3.1.4.4 3.2 水解 O-糖基化合物 3.2.1 糖苷酸 α-淀粉酶 3.2.1.1 β-淀粉酶 3.2.1.2 葡糖糖化酶 3.2.1.3 纤维素酶 3.2.1.4 聚半乳糖醛酸酶 3.2.1.15 溶菌酶 3.2.1.17 α-D-糖苷酶(麦芽糖酶) 3.2.1.20 β-D-糖苷酶 3.2.1.21 α-D-半乳糖苷酶 3.2.1.22 β-D-半乳糖苷酶(乳糖酶) 3.2.1.23 β-呋喃果糖苷酶(转化酶或蔗糖 酶) 3.2.1.26 1,3-β-D-木聚糖酶 3.2.1.32 α-L-鼠李糖苷酶 3.2.1.40 支链淀粉酶 3.2.1.41 外切聚半乳糖醛酸酶 3.2.3.67 3.2.3 水解 S-糖基化合物 葡糖硫苷酶(黑芥子硫苷酸酶) 3.2.3.1 3.4 肽酶
- 8 - 1.14 作用于一对供体 一元酚单加氧酶(多酚氧化酶) 1.14.18.1 2 转移酶 2.7 转移磷酸 2.7.1 受体为 OH 己糖激酶 2.7.1.1 甘油激酶 2.7.1.30 丙酮酸激酶 2.7.1.40 2.7.3 受体为 N-基 肌酸激酶 2.7.3.2 3 水解酶 3.1 切断酯键 3.1.1 羧酸酯水解酶 羧酸酯酶 3.1.1.1 三酰甘油酯酶 3.1.1.3 磷酸酯酶A2 3.1.1.4 乙酰胆碱酯酶 3.1.1.7 果胶甲酯酶 3.1.1.11 磷酸酯酶A1 3.1.1.32 3.1.3 磷酸单酯水解酶 碱性磷酸酯酶 3.1.3.1 3.1.4 磷酸双酯水解酶 磷脂酶 C 3.1.4.2 磷脂酶 D 3.1.4.4 3.2 水解 O-糖基化合物 3.2.1 糖苷酸 α-淀粉酶 3.2.1.1 β-淀粉酶 3.2.1.2 葡糖糖化酶 3.2.1.3 纤维素酶 3.2.1.4 聚半乳糖醛酸酶 3.2.1.15 溶菌酶 3.2.1.17 α-D-糖苷酶(麦芽糖酶) 3.2.1.20 β-D-糖苷酶 3.2.1.21 α-D-半乳糖苷酶 3.2.1.22 β-D-半乳糖苷酶(乳糖酶) 3.2.1.23 β-呋喃果糖苷酶(转化酶或蔗糖 酶) 3.2.1.26 1,3-β-D-木聚糖酶 3.2.1.32 α-L-鼠李糖苷酶 3.2.1.40 支链淀粉酶 3.2.1.41 外切聚半乳糖醛酸酶 3.2.3.67 3.2.3 水解 S-糖基化合物 葡糖硫苷酶(黑芥子硫苷酸酶) 3.2.3.1 3.4 肽酶

- 9 - 3.4.21 丝氨酸肽键内切酶 微生物丝氨酸肽键内切酶如枯草杆 菌蛋白酶 3.4.21.62 3.4.23 天冬氨酸肽键内切酶 凝乳酶 3.4.23.4 3.4.24 金属肽键内切酶 嗜热菌蛋白酶 3.4.24.27 3.5 作用于除肽键外的 C-N 键 3.5.2 环内酰胺中 肌酸酐酶 3.5.2.10 4 裂解酶 4.2 C-O-裂解酶 4.2.2 作用于多糖 果胶酸裂解酶 4.2.2.2 外切聚半乳糖醛酸裂解酶 4.2.2.9 果胶裂解酶 4.2.2.10 5 异构酶 5.3 分子内氧化还原酶 5.3.1 醛糖和酮糖间的互变 木糖异构酶 5.3.1.5 葡萄糖-6-磷酸异构酶 5.3.1.9 四、生物体中的酶 自然界所有的生物体中都含有许多种类的酶,一些特殊的酶仅存在于细 胞内的一类细胞器中。细胞核中含有的酶主要涉及到核酸的生物合成和水解 降解,线粒体中含有与氧化磷酸化和生成 ATP 有关的氧化还原酶。细胞中的 许多酶常常在一个连续的反应链中起作用。酶在生物体内是非均匀分布的, 特定的器官含有特定的酶,在植物组织中这些酶随着生长、发育,其数量将 会发生变化,有的甚至十分显著。 在完整细胞内,许多多酶体系具有自我调节的能力,酶的活力一般是通 过以下几种方式控制:隔离分布在亚细胞膜内,被细胞器控制;酶结合于膜 或细胞壁上;酶作用的底物结合于膜或细胞壁;酶与底物分离。此外,还可 通过依靠酶原的生物合成和生理上重要的内源酶抑制剂控制。 一旦组织受损将使酶与底物接近,导致食品在色泽、质地、风味和营养 上的改变。因此,热处理、低温保藏和酶抑制剂的使用均有利于稳定产品质 量。 食品加工的生物材料中含有数以百计的不同种类的酶,这些酶对于原料 的生长、成熟、加工和保藏等均起着重要的作用。表 6-3 列举了一些食品原 料中几种酶的相对含量。可见不同的植物中各种酶的含量差异较大
- 9 - 3.4.21 丝氨酸肽键内切酶 微生物丝氨酸肽键内切酶如枯草杆 菌蛋白酶 3.4.21.62 3.4.23 天冬氨酸肽键内切酶 凝乳酶 3.4.23.4 3.4.24 金属肽键内切酶 嗜热菌蛋白酶 3.4.24.27 3.5 作用于除肽键外的 C-N 键 3.5.2 环内酰胺中 肌酸酐酶 3.5.2.10 4 裂解酶 4.2 C-O-裂解酶 4.2.2 作用于多糖 果胶酸裂解酶 4.2.2.2 外切聚半乳糖醛酸裂解酶 4.2.2.9 果胶裂解酶 4.2.2.10 5 异构酶 5.3 分子内氧化还原酶 5.3.1 醛糖和酮糖间的互变 木糖异构酶 5.3.1.5 葡萄糖-6-磷酸异构酶 5.3.1.9 四、生物体中的酶 自然界所有的生物体中都含有许多种类的酶,一些特殊的酶仅存在于细 胞内的一类细胞器中。细胞核中含有的酶主要涉及到核酸的生物合成和水解 降解,线粒体中含有与氧化磷酸化和生成 ATP 有关的氧化还原酶。细胞中的 许多酶常常在一个连续的反应链中起作用。酶在生物体内是非均匀分布的, 特定的器官含有特定的酶,在植物组织中这些酶随着生长、发育,其数量将 会发生变化,有的甚至十分显著。 在完整细胞内,许多多酶体系具有自我调节的能力,酶的活力一般是通 过以下几种方式控制:隔离分布在亚细胞膜内,被细胞器控制;酶结合于膜 或细胞壁上;酶作用的底物结合于膜或细胞壁;酶与底物分离。此外,还可 通过依靠酶原的生物合成和生理上重要的内源酶抑制剂控制。 一旦组织受损将使酶与底物接近,导致食品在色泽、质地、风味和营养 上的改变。因此,热处理、低温保藏和酶抑制剂的使用均有利于稳定产品质 量。 食品加工的生物材料中含有数以百计的不同种类的酶,这些酶对于原料 的生长、成熟、加工和保藏等均起着重要的作用。表 6-3 列举了一些食品原 料中几种酶的相对含量。可见不同的植物中各种酶的含量差异较大

- 10 - 表 6-3 不同原料中几种酶的相对含量 酶 来源 相对含量(%) 聚半乳糖醛酸酶 番茄 1.0 鳄梨 0.065 欧楂(Medlar) 0.027 梨 0.016 菠萝 0.024 胡萝卜 0 葡萄 0 脂肪氧合酶 大豆 1.00 摩尔拉豆(Urd bean) 0.64 绿豆 0.47 碗豆 0.32 小麦 0.02 花生 0.01 过氧化物酶 青刀豆 1.00 豆荚 0.72 菜豆 0.62 菠菜 0.32 利马豆 0.15 多酚氧化酶是植物中最受注意的一种酶,在葡萄、洋李、无花果、枣、 茶叶和咖啡豆中含量很高,它在这些果实中起着人们期望的作用。另外,多 酚氧化酶在桃、苹果、香蕉、荔枝、马铃薯、莲藕和莴苣中的含量也是相当 高,然而它对这些果实起着不需宜的作用,易引起褐变,造成变质和腐烂, 对于新鲜果实的保藏带来极大困难。胡椒中不存在多酚氧化酶。 酶在食品中的活力与其生长环境和成熟度有关,从而给生产质量的稳定 性带来诸多不便。幸运的是,酶变性的速率依然遵循一级反应动力学,因此, 酶失活到一定程度所需的时间与酶的浓度无关。然而不同起始浓度的酶失活, 即使将酶失活到相同的百分数,但剩余的绝对酶活仍然是不同的。对于起始 浓度高的酶,完全失活则需要较长时间。食品原料中的同功酶往往具有不同 的热稳定性。 五、酶的分离纯化与活力测定 1、酶的分离纯化
- 10 - 表 6-3 不同原料中几种酶的相对含量 酶 来源 相对含量(%) 聚半乳糖醛酸酶 番茄 1.0 鳄梨 0.065 欧楂(Medlar) 0.027 梨 0.016 菠萝 0.024 胡萝卜 0 葡萄 0 脂肪氧合酶 大豆 1.00 摩尔拉豆(Urd bean) 0.64 绿豆 0.47 碗豆 0.32 小麦 0.02 花生 0.01 过氧化物酶 青刀豆 1.00 豆荚 0.72 菜豆 0.62 菠菜 0.32 利马豆 0.15 多酚氧化酶是植物中最受注意的一种酶,在葡萄、洋李、无花果、枣、 茶叶和咖啡豆中含量很高,它在这些果实中起着人们期望的作用。另外,多 酚氧化酶在桃、苹果、香蕉、荔枝、马铃薯、莲藕和莴苣中的含量也是相当 高,然而它对这些果实起着不需宜的作用,易引起褐变,造成变质和腐烂, 对于新鲜果实的保藏带来极大困难。胡椒中不存在多酚氧化酶。 酶在食品中的活力与其生长环境和成熟度有关,从而给生产质量的稳定 性带来诸多不便。幸运的是,酶变性的速率依然遵循一级反应动力学,因此, 酶失活到一定程度所需的时间与酶的浓度无关。然而不同起始浓度的酶失活, 即使将酶失活到相同的百分数,但剩余的绝对酶活仍然是不同的。对于起始 浓度高的酶,完全失活则需要较长时间。食品原料中的同功酶往往具有不同 的热稳定性。 五、酶的分离纯化与活力测定 1、酶的分离纯化