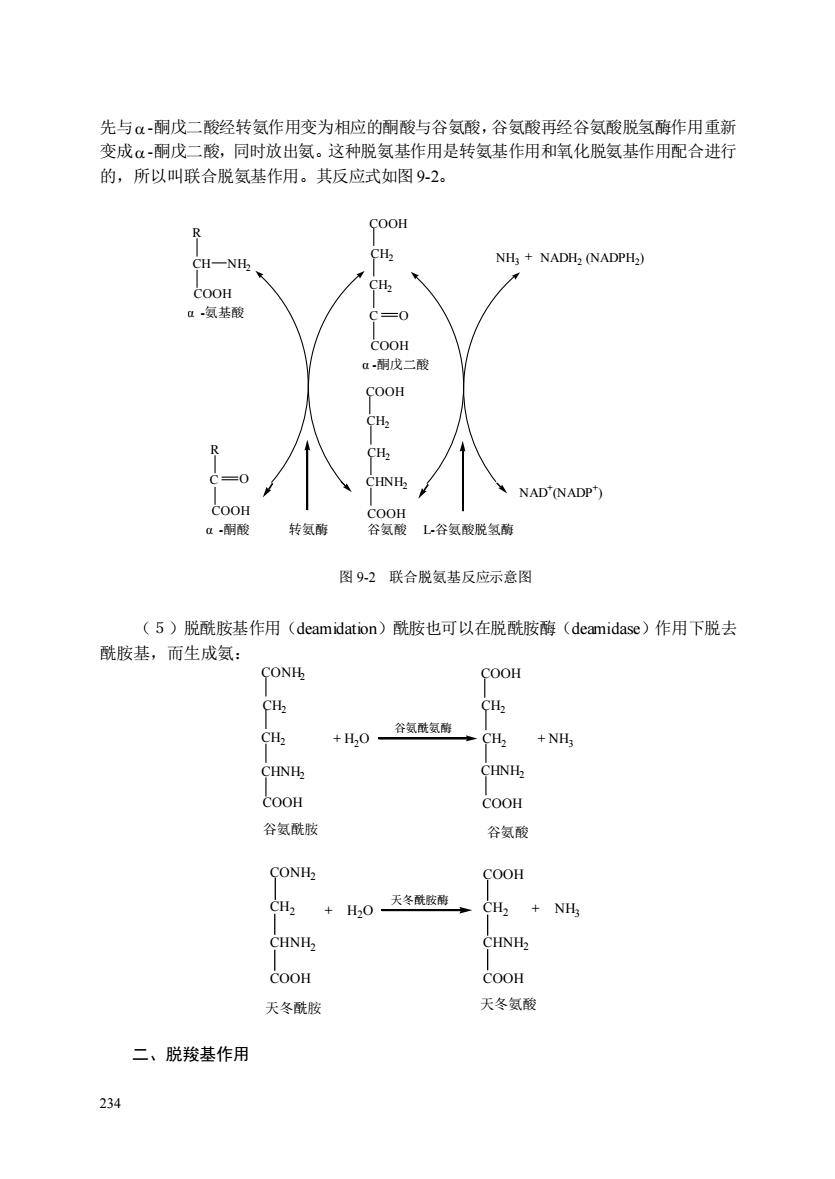
先与α-酮戊二酸经转氨作用变为相应的酮酸与谷氨酸,谷氨酸再经谷氨酸脱氢酶作用重新 变成α-酮戊二酸,同时放出氨。这种脱氨基作用是转氨基作用和氧化脱氨基作用配合进行 的,所以叫联合脱氨基作用。其反应式如图92。 NH:+NADH,(NADPH) 氨基酸 OH COOH R CH 0 NAD'(NADP COOH 转氨酶 谷酸L谷氨酸脱氢酶 图92联合脱氨基反应示意图 (5)脱酰胺基作用(deamidation)酰胺也可以在脱酰胺酶(deamidase)作用下脱去 酰胺基,而生成氨: CONH COOH CH2 +山,0一谷氨酰氨酶 +NH; CHNH CHNH COOH COOH 谷氨酰胺 谷氨酸 CONH C +H20天冬酰胶酶 CH2 NH3 COOH COOH 天冬酰胺 天冬氨酸 二、脱羧基作用 234
234 先与 -酮戊二酸经转氨作用变为相应的酮酸与谷氨酸,谷氨酸再经谷氨酸脱氢酶作用重新 变成 -酮戊二酸,同时放出氨。这种脱氨基作用是转氨基作用和氧化脱氨基作用配合进行 的,所以叫联合脱氨基作用。其反应式如图 9-2。 COOH CH2 CH2 C COOH COOH CH2 CH2 CHNH2 COOH 谷氨酸 NH3 R CH COOH NH2 O R COOH C O α -氨基酸 α -酮酸 转氨酶 α -酮戊二酸 L-谷氨酸脱氢酶 + NADH2 (NADPH2 ) NAD + (NADP + ) 图 9-2 联合脱氨基反应示意图 (5)脱酰胺基作用(deamidation)酰胺也可以在脱酰胺酶(deamidase)作用下脱去 酰胺基,而生成氨: CONH2 CH2 CH2 CHNH2 COOH + H2O 谷氨酰氨酶 COOH CH2 CH2 CHNH2 COOH 谷氨酰胺 谷氨酸 + NH3 二、脱羧基作用 CONH2 CH2 CHNH2 COOH + H2O 天冬酰胺酶 COOH CH2 CHNH2 COOH 天冬酰胺 天冬氨酸 + NH3
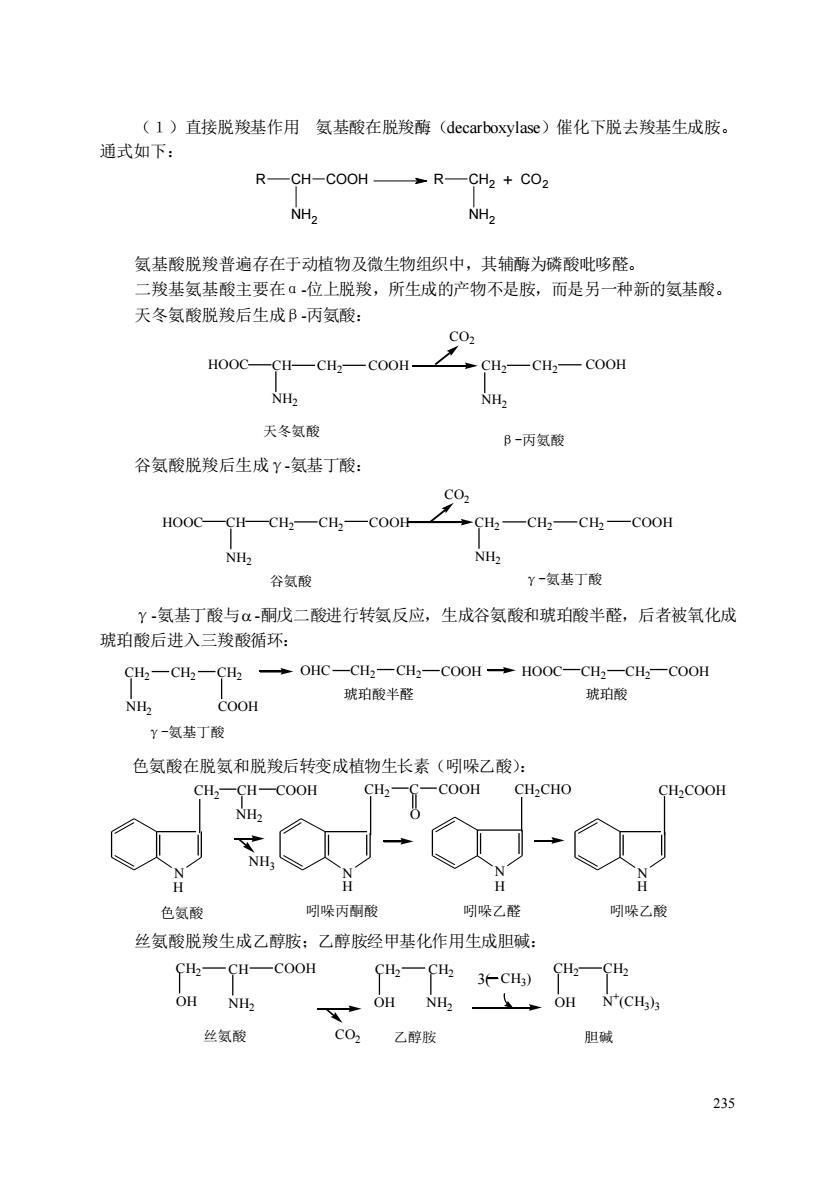
(1)直接脱羧基作用氨基酸在脱羧酶(decarboxylase)催化下脱去羧基生成胺 通式如下: R—CH-COOH→R—CH2+CO NH2 NH 氨基酸脱羧普遍存在于动植物及微生物组织中,其辅酶为磷酸吡哆醛。 二羧基氨基酸主要在ā位上脱羧,所生成的产物不是胺,而是另一种新的氨基酸。 天冬氨酸脱羧后生成B丙氨酸: HOOC—CH—CH2—COOH →CH—CH2一COH NH2 NH2 天冬氨酸 B-丙氨酸 谷氨酸脱羧后生成Y-氨基丁酸: oa0一a4-0-oon-2 CH2—CH—Ch—COOH NHz NHz 谷氨酸 Y-氨基丁酸 Y-氨基丁酸与α-酮成二酸进行转氨反应,生成谷氨酸和琥珀酸半醛,后者被氧化成 琥珀酸后进入三羧酸循环: SH-C4一H,→oHC-CH-CH-C00H→Ho0C一CH-CHh一co0H NH COOH 琥珀酸半醛 琥珀酸 y-氨基丁酸 色氨酸在脱氨和脱羧后转变成植物生长素(吲哚乙酸): CH,一CH-COOH CH2COOH NH2 NH, 色氨酸 明哚丙解酸 吲吼哚乙醛 吲哚乙酸 丝氨酸脱羧生成乙醇胺:乙醇胺经甲基化作用生成胆碱: FH一CH一COoH 4一H3C) H—4 OH NH2 OH OH N(CH)s 丝氨酸 CO2乙醇胺 胆碱 235
235 (1)直接脱羧基作用 氨基酸在脱羧酶(decarboxylase)催化下脱去羧基生成胺。 通式如下: 氨基酸脱羧普遍存在于动植物及微生物组织中,其辅酶为磷酸吡哆醛。 二羧基氨基酸主要在α-位上脱羧,所生成的产物不是胺,而是另一种新的氨基酸。 天冬氨酸脱羧后生成β-丙氨酸: HOOC CH CH2 COOH CO2 CH2 COOH NH2 NH2 天冬氨酸 β-丙氨酸 CH2 谷氨酸脱羧后生成γ-氨基丁酸: HOOC CH CH2 COOH CO2 CH2 CH2 COOH NH2 NH2 CH2 谷氨酸 γ-氨基丁酸 CH2 γ-氨基丁酸与 -酮戊二酸进行转氨反应,生成谷氨酸和琥珀酸半醛,后者被氧化成 琥珀酸后进入三羧酸循环: CH2 NH2 CH2 CH2 COOH OHC CH2 CH2 COOH HOOC CH2 CH2 COOH γ-氨基丁酸 琥珀酸半醛 琥珀酸 色氨酸在脱氨和脱羧后转变成植物生长素(吲哚乙酸): N N N CH2 CH COOH NH2 NH3 CH2 C O COOH CH2CHO N H H H H 色氨酸 吲哚丙酮酸 吲哚乙醛 吲哚乙酸 CH2COOH 丝氨酸脱羧生成乙醇胺;乙醇胺经甲基化作用生成胆碱: CO2 CH2 CH COOH NH2 CH2 CH2 CH2 CH2 OH NH2 OH N + (CH3 ) OH 3 丝氨酸 乙醇胺 胆碱 3( CH3 ) CH COOH NH2 R CH2 NH2 R + CO2

乙醇胺和胆碱分别是脑磷脂和卵磷脂的成分。 某些胺的氨基酸前体见表94。 表94某些胺的氨基酸前体 氨基酸 胺 结构式 丝氨酸 乙醇胺 CH-CHNH 微氨酸 丁胺 a-og4 异亮氨酸 异戊胺 -M 甲硫氨酸 甲硫基丙胶 CH:-CH-CHNHE SCH: 赖氨酸 尸胺 CH-(CH2)-CHNH N 鸟氨酸 腐胺 C一Cep一CH-NH NH 精氨酸 精胺 H2N(CH2)NH(CH2NH (CH2NH: 亚桔胺 HN(CH)NH(CH2pNH 苯丙氨酸 苯乙胺 CHCH.NH 酪氨酸 酪胺 CH-CH.NH: 色氨酸 CH2-CH2NHz 这些胺类在植物体内进一步转化所形成的产物都具有一定的生理作用。胺可经氨氧化 酶氧化成醛和氨:醛经脱氢酶作用氧化成脂肪酸:脂肪酸经-氧化生成乙酰辅酶A而进入 三羧酸循环彻底氧化: +0、+1L0 氢格RCOOH一B氧化 +1204 RCHO 酸 TCA环 C02+H,0 (2)羟化脱羟基作用(ydroxylation)酪氨酸在酪氨酸酶(tyrosinase)催化下发生 羟化而生成3,4二羟苯丙氨酸,简称多巴,后者可脱羧生成3,4二羟苯乙胺,简称多巴 胺: 236
236 乙醇胺和胆碱分别是脑磷脂和卵磷脂的成分。 某些胺的氨基酸前体见表 9-4。 表 9-4 某些胺的氨基酸前体 氨 基 酸 胺 结 构 式 丝氨酸 缬氨酸 异亮氨酸 甲硫氨酸 赖氨酸 鸟氨酸 精氨酸 苯丙氨酸 酪氨酸 色氨酸 乙醇胺 丁 胺 异戊胺 甲硫基丙胺 尸 胺 腐 胺 精 胺 亚精胺 苯乙胺 酪 胺 色 胺 CH2—CH2NH2 ∣ OH CH3 CH CH2NH2 CH3 CH3 CH CH2NH2 CH3 CH2 CH2—CH2—CH2NH2 ∣ SCH3 CH2—(CH2)3—CH2NH2 ∣ NH2 CH2—(CH2)2—CH2NH2 ∣ NH2 H2N (CH2)3NH (CH2)4NH (CH2)3NH2 H2N (CH2)4NH (CH2)3NH2 CH2 CH2NH2 HO CH2 CH2NH2 N H CH2 CH2NH2 这些胺类在植物体内进一步转化所形成的产物都具有一定的生理作用。胺可经氨氧化 酶氧化成醛和氨;醛经脱氢酶作用氧化成脂肪酸;脂肪酸经 -氧化生成乙酰辅酶 A 而进入 三羧酸循环彻底氧化: RCH2NH2 +O2 + H2O +1/2O2 RCHO RCOOH CO2 + H2O TCA环 β -氧化 胺氧化酶 醛脱氢酶 胺 醛 酸 (2)羟化脱羟基作用(hydroxylation)酪氨酸在酪氨酸酶(tyrosinase)催化下发生 羟化而生成 3,4-二羟苯丙氨酸,简称多巴,后者可脱羧生成 3,4-二羟苯乙胺,简称多巴 胺:
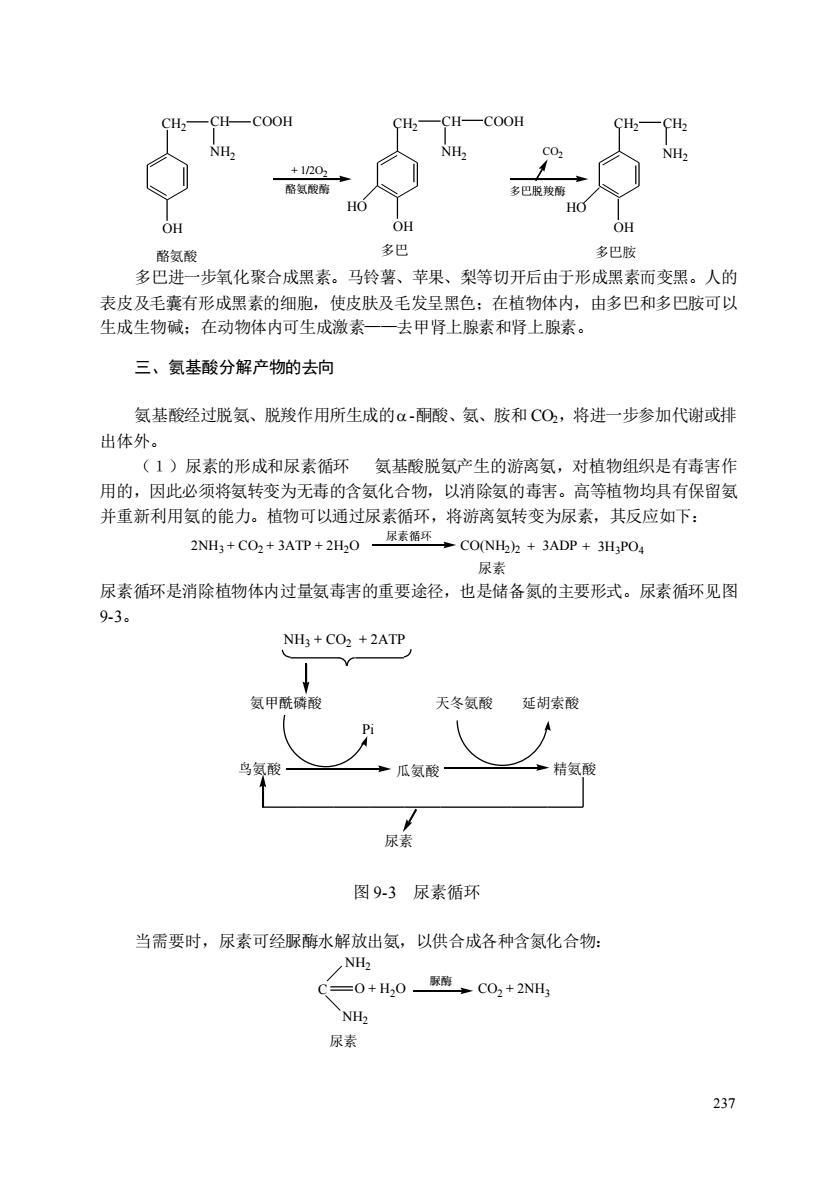
CH>- -CH—COOH CH-COOH NH +1/2G 氨酸 多巴脱 OH 多化聚合成素。马、平果、梨等切开后由于形熙素而变。人的 多巴 名巴胺 表皮及毛囊有形成黑素的细胞,使皮肤及毛发呈黑色:在植物体内,由多巴和多巴胺可以 生成生物碱:在动物体内可生成激素 去甲肾上腺素和肾上腺素。 三、氨基酸分解产物的去向 氢基酸经过脱氨、脱羧作用所生成的α-酮酸、氢、胺和CO2,将进一步参加代谢或排 出体外。 (1)尿素的形成和尿素循环氨基酸脱氨产生的游离氨,对植物组织是有毒害作 用的,因此必须将氨转变为无毒的含氨化合物,以消除氨的毒害。高等植物均具有保留氨 并重新利用氨的能力。植物可以通过尿素循环,将游离氨转变为尿素,其反应如下: 2NH3 +CO2+3ATP+2H20 →CO(NH2)+3ADP+3H,PO 尿动 尿素循环是消除植物体内过量氨毒害的重要途径,也是储备氮的主要形式。尿素循环见图 9-3。 NH3 +CO2 +2ATP 氨甲酰酸 天冬氨酸延胡索酸 ”瓜氨酸 精氨酸 尿素 图93尿素循环 当需要时,尿素可经脲酶水解放出氨,以供合成各种含氮化合物: NH2 C二0+H,0、CO2+2NH) NH 尿素 231
237 CH2 CH COOH NH2 CH2 CH COOH NH2 CH2 CH2 HO HO OH OH OH + 1/2O2 CO2 酪氨酸酶 多巴脱羧酶 NH2 酪氨酸 多巴 多巴胺 多巴进一步氧化聚合成黑素。马铃薯、苹果、梨等切开后由于形成黑素而变黑。人的 表皮及毛囊有形成黑素的细胞,使皮肤及毛发呈黑色;在植物体内,由多巴和多巴胺可以 生成生物碱;在动物体内可生成激素——去甲肾上腺素和肾上腺素。 三、氨基酸分解产物的去向 氨基酸经过脱氨、脱羧作用所生成的 -酮酸、氨、胺和 CO2,将进一步参加代谢或排 出体外。 (1)尿素的形成和尿素循环 氨基酸脱氨产生的游离氨,对植物组织是有毒害作 用的,因此必须将氨转变为无毒的含氨化合物,以消除氨的毒害。高等植物均具有保留氨 并重新利用氨的能力。植物可以通过尿素循环,将游离氨转变为尿素,其反应如下: 2NH3 + CO2 + 3ATP + 2H2O 尿素循环 尿素 CO(NH2 )2 + 3ADP + 3H3PO4 尿素循环是消除植物体内过量氨毒害的重要途径,也是储备氮的主要形式。尿素循环见图 9-3。 NH3 + CO2 + 2ATP 氨甲酰磷酸 天冬氨酸 延胡索酸 鸟氨酸 瓜氨酸 精氨酸 尿素 Pi 图 9-3 尿素循环 当需要时,尿素可经脲酶水解放出氨,以供合成各种含氮化合物: NH2 CO2 + 2NH3 脲酶 NH2 尿素 C O + H2O

(2)合成其他含氮化合物植物可将游离氨转变成氨基酸、酰胺、有机酸铵盐及氨 甲酰磷酸等含氮化合物,以消除氨的毒害和储存以备再度利用。 如果植物组织中含有足够的碳水化合物,氨可与碳水化合物转化的酮酸进行氨基化反 应,重新生成氨基酸。 在植物体中,消除氨的毒害作用的途径主要是生成酰胺化合物。天冬酰胺和谷氨酰胺 是动植物共有的储氮形式。当体内酮酸增多时,再经转氨作用放出氨,生成氨基酸。 有些植物组织中含有大量的有机酸,如秋海棠、酸模、大黄等含有柠檬酸、草酰乙酸、 苹果酸等,氨便与有机酸形成有机酸的铵盐 氨与CO2(来自三羧酸循环)在AP参与下,经酶催化形成氨基甲酰磷酸: NH.CO:ADP 氨甲酰磷酸是合成嘧啶、瓜氨酸、精氨酸和尿素的主要代谢物,也是植物和微生物保 存氮的重要方式。 (3)氨基酸碳架的氧化氨基酸脱氨后余下的碳架要进一步转化,最后生成各种有 机酸。各种氨基酸的代谢产物列于表95中。 表95氨基酸代谢的终产物 氨基酸 终产物 丙氨酸、丝氨酸、半秋酸、酰氨酸、甘氨酸、苏氨酸 丙制酸 亮氨酸 乙酰辅南A 苯丙氨酸、酪氨酸、亮酸、赖氨酸、色氨酸 乙酰乙酸(或乙酰乙酰辅酶A) 甲硫氨酸、异亮氨酸。氨酸 虢珀酰辅酶A 苯丙氨酸、酪氨酸 延胡索酸 精氨酸、酌氨酸、组氨酸、谷氨酰贼、谷氨酸 a酮成二酸 天冬氯酸、天冬酰胺 草酰乙酸 从表95中氨基酸分解代谢的终产物可以看出,除乙酰乙酰辅酶A外,均是糖酵解和 三羧酸循环的中间产物,而乙酰乙酰辅酶A也可以分解为乙酰辅酶A,所以这些中间产物 最后均可通过三羧酸循环而氧化分解,如图94所示。 (4)转变成糖和脂类氨基酸脱氨后的碳架,根据有机体代谢的需要,即可经过三 羧酸循环彻底氧化。生成ATP供机体能量的需要,又可以转变成糖和脂肪。 凡是形成丙酮酸、α-酮成一酸、琥珀酸、草酰乙酸的氨基酸都称为生糖氨基酸,因为 这些有机酸经过转变都能生成葡萄糖。 凡能生成乙酰辅酶A和乙酰乙酰辅酶A的氨基酸均能通过乙酰辅酶A转变成脂肪。 有些氨基酸(如苯丙氨酸和酪氨酸)既可生成脂肪又可生成糖。 238
238 (2)合成其他含氮化合物 植物可将游离氨转变成氨基酸、酰胺、有机酸铵盐及氨 甲酰磷酸等含氮化合物,以消除氨的毒害和储存以备再度利用。 如果植物组织中含有足够的碳水化合物,氨可与碳水化合物转化的酮酸进行氨基化反 应,重新生成氨基酸。 在植物体中,消除氨的毒害作用的途径主要是生成酰胺化合物。天冬酰胺和谷氨酰胺 是动植物共有的储氮形式。当体内酮酸增多时,再经转氨作用放出氨,生成氨基酸。 有些植物组织中含有大量的有机酸,如秋海棠、酸模、大黄等含有柠檬酸、草酰乙酸、 苹果酸等,氨便与有机酸形成有机酸的铵盐。 氨与 CO2(来自三羧酸循环)在 ATP 参与下,经酶催化形成氨基甲酰磷酸: NH3 CO2 ATP H2N C O O P ADP Mg 2+ + + + 氨甲酰磷酸是合成嘧啶、瓜氨酸、精氨酸和尿素的主要代谢物,也是植物和微生物保 存氮的重要方式。 (3)氨基酸碳架的氧化 氨基酸脱氨后余下的碳架要进一步转化,最后生成各种有 机酸。各种氨基酸的代谢产物列于表 9-5 中。 表 9-5 氨基酸代谢的终产物 氨 基 酸 终 产 物 丙氨酸、丝氨酸、半胱氨酸、胱氨酸、甘氨酸、苏氨酸 亮氨酸 苯丙氨酸、酪氨酸、亮氨酸、赖氨酸、色氨酸 甲硫氨酸、异亮氨酸、缬氨酸 苯丙氨酸、酪氨酸 精氨酸、脯氨酸、组氨酸、谷氨酰胺、谷氨酸 天冬氨酸、天冬酰胺 丙酮酸 乙酰辅酶 A 乙酰乙酸(或乙酰乙酰-辅酶 A) 琥珀酰辅酶 A 延胡索酸 -酮戊二酸 草酰乙酸 从表 9-5 中氨基酸分解代谢的终产物可以看出,除乙酰乙酰辅酶 A 外,均是糖酵解和 三羧酸循环的中间产物,而乙酰乙酰辅酶 A 也可以分解为乙酰辅酶 A,所以这些中间产物 最后均可通过三羧酸循环而氧化分解,如图 9-4 所示。 (4)转变成糖和脂类 氨基酸脱氨后的碳架,根据有机体代谢的需要,即可经过三 羧酸循环彻底氧化。生成 ATP 供机体能量的需要,又可以转变成糖和脂肪。 凡是形成丙酮酸、 -酮戊一酸、琥珀酸、草酰乙酸的氨基酸都称为生糖氨基酸,因为 这些有机酸经过转变都能生成葡萄糖。 凡能生成乙酰辅酶 A 和乙酰乙酰辅酶 A 的氨基酸均能通过乙酰辅酶 A 转变成脂肪。 有些氨基酸(如苯丙氨酸和酪氨酸)既可生成脂肪又可生成糖