NANA Gal Gle CH OH OH CH CH HN o.O CH OH CH CH OH CHOH R CHOH CH,OH 1 galactocerebroside 2 Gu ganglioside 3 sialic acid (NANA) 图4-6糖脂的结构1.半乳糖脑苷脂,2.GM1神经节苷脂,3.唾液酸 (三)、胆固酵 胆固醇(图4-7)仅存在真核细胞膜上,含量一般不超过膜脂的113,植物细胞膜中含量较 少,其功能是提高脂双层的力学稳定性,调节脂双层流动性,降低水溶性物质的通透性。如: 在缺少胆固醇培养基中,不能合成胆固醇的突变细胞株很快发生自溶
图 4-6 糖脂的结构 1. 半乳糖脑苷脂,2. GM1 神经节苷脂,3. 唾液酸 (三)、胆固醇 胆固醇(图 4-7)仅存在真核细胞膜上,含量一般不超过膜脂的 1/3,植物细胞膜中含量较 少,其功能是提高脂双层的力学稳定性,调节脂双层流动性,降低水溶性物质的通透性。如: 在缺少胆固醇培养基中,不能合成胆固醇的突变细胞株很快发生自溶
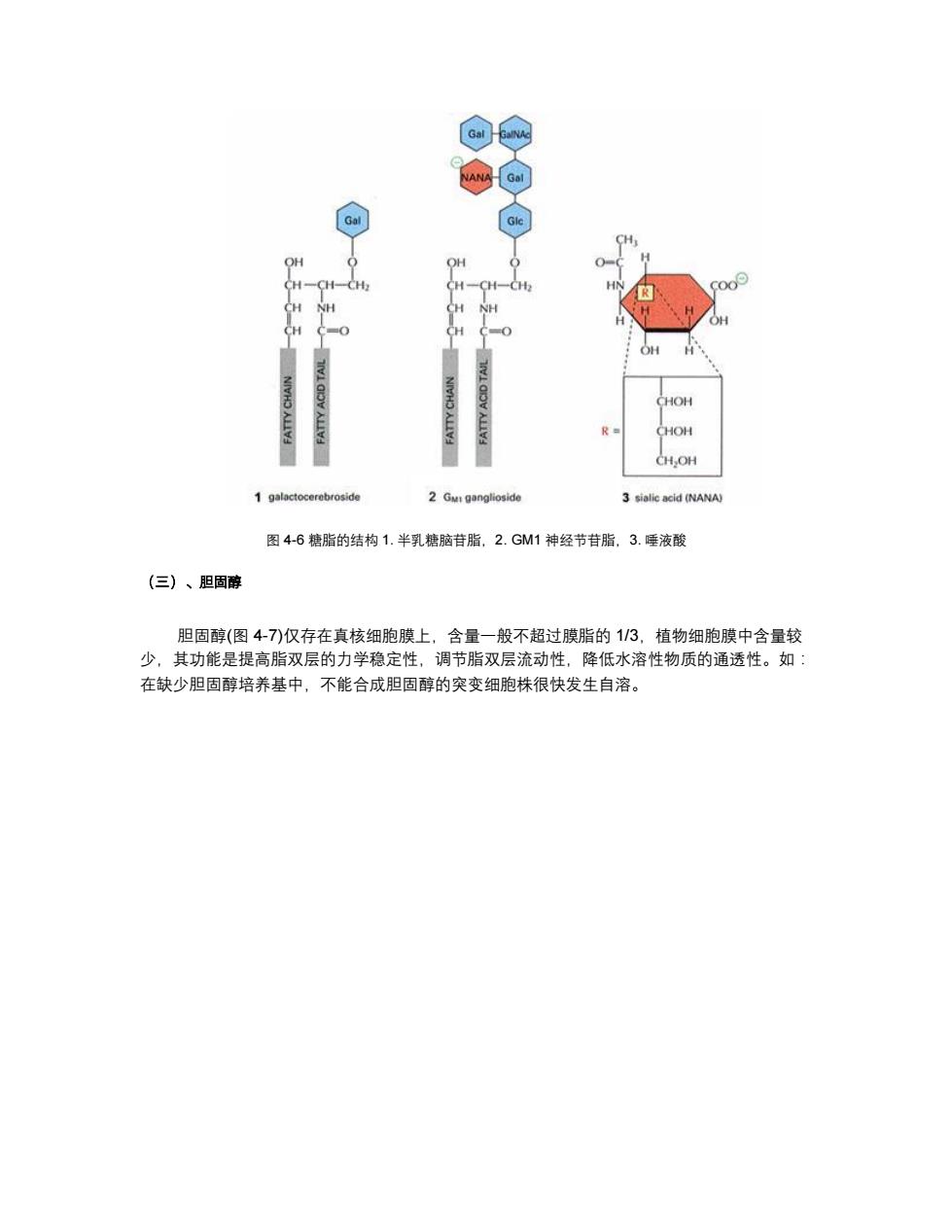
(a) 18 (b) 2 17 CH3 11 D16 27 19 5 25HC-CHg 2 10 9 8 3 5 B 7 24CH2 4 6 23CH2 22CH2 HC20 CHa 18 21 H3l 19 3 2 9 10 A 8 5 4 6 HO Hydrophilic Hydrophobic region region 图4-7胆固醇 (四)、脂质体 脂质体((liposome)是一种人工膜。在水中磷脂分子亲水头部插入水中,疏水尾部伸向空 气,搅动后形成双层脂分子的球形脂质体,直径25~1000nm不等。脂质体可用于转基因,或 制备的药物,利用脂质体可以和细胞膜融合的特点,将药物送入细胞内部
图 4-7 胆固醇 (四)、脂质体 脂质体(liposome)是一种人工膜。在水中磷脂分子亲水头部插入水中,疏水尾部伸向空 气,搅动后形成双层脂分子的球形脂质体,直径 25~1000nm 不等。脂质体可用于转基因,或 制备的药物,利用脂质体可以和细胞膜融合的特点,将药物送入细胞内部
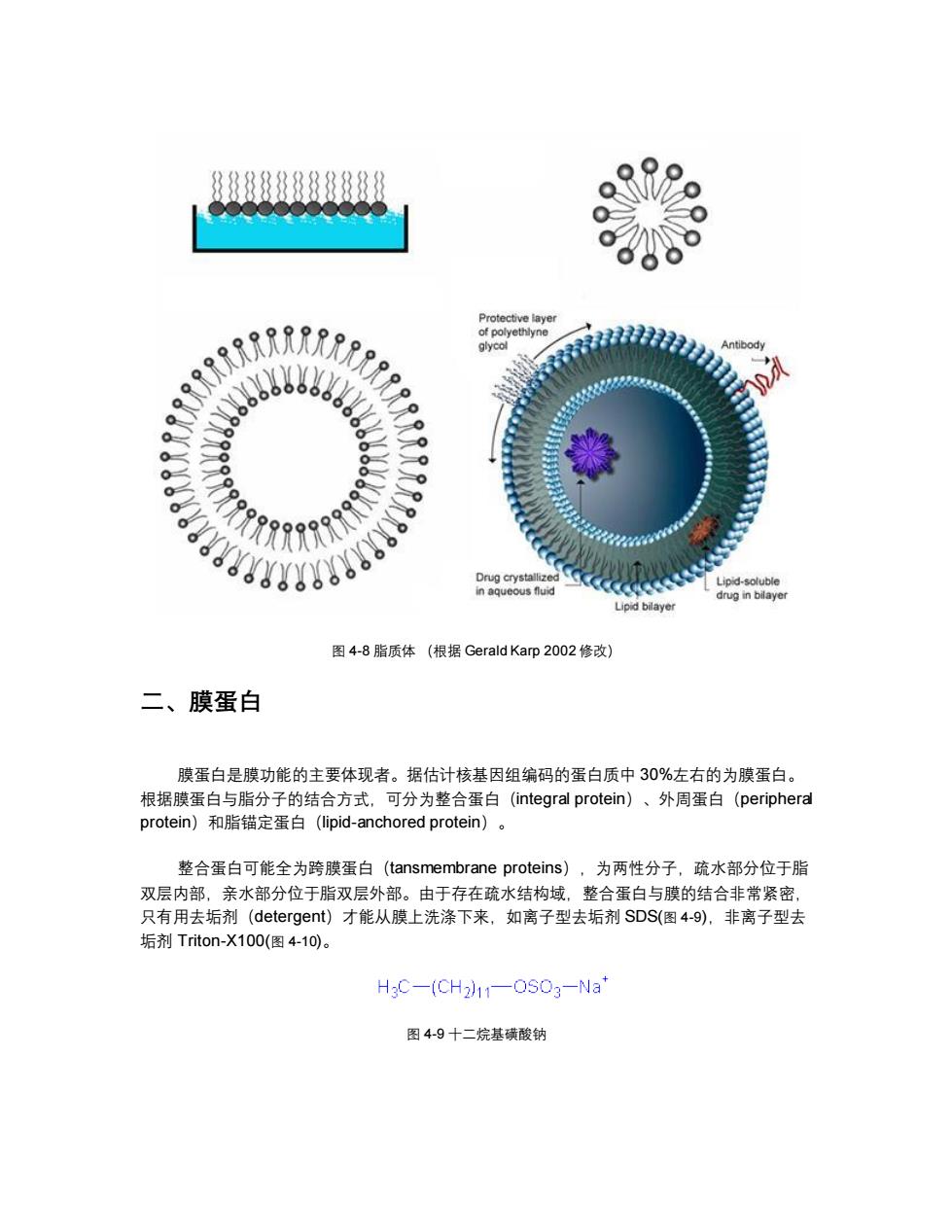
8888888888 Protective layer of polyethlyne glycol 322 tibody 0 Drug crystallized Lipid-soluble in aqueous fluid drug in bilayer Lipid bilayer 图4-8脂质体(根据Gerald Karp2002修改) 二、膜蛋白 膜蛋白是膜功能的主要体现者。据估计核基因组编码的蛋白质中30%左右的为膜蛋白。 根据膜蛋白与脂分子的结合方式,可分为整合蛋白(integral protein)、外周蛋白(peripheral protein)和脂锚定蛋白(lipid-anchored protein)o 整合蛋白可能全为跨膜蛋白(tansmembrane proteins),为两性分子,疏水部分位于脂 双层内部,亲水部分位于脂双层外部。由于存在疏水结构域,整合蛋白与膜的结合非常紧密, 只有用去垢剂(detergent)才能从膜上洗涤下来,如离子型去垢剂SDS(图4-9),非离子型去 垢剂Triton-X100(图4-10)。 H3C-(CH2)11-OSO3-Na* 图4-9十二烷基磺酸钠
图 4-8 脂质体 (根据 Gerald Karp 2002 修改) 二、膜蛋白 膜蛋白是膜功能的主要体现者。据估计核基因组编码的蛋白质中 30%左右的为膜蛋白。 根据膜蛋白与脂分子的结合方式,可分为整合蛋白(integral protein)、外周蛋白(peripheral protein)和脂锚定蛋白(lipid-anchored protein)。 整合蛋白可能全为跨膜蛋白(tansmembrane proteins),为两性分子,疏水部分位于脂 双层内部,亲水部分位于脂双层外部。由于存在疏水结构域,整合蛋白与膜的结合非常紧密, 只有用去垢剂(detergent)才能从膜上洗涤下来,如离子型去垢剂 SDS(图 4-9),非离子型去 垢剂 Triton-X100(图 4-10)。 图 4-9 十二烷基磺酸钠

CH3 CH3 (0-CH2-CH210-0H CH3 图4-10 Triton-X100 整合蛋白的跨膜结构域可以是1至多个疏水的α螺旋,形成亲水通道的整合蛋白跨膜区 域有两种组成形式,一是由多个两性α螺旋组成亲水通道;而是由两性B折叠组成亲水通道 (图4-11)。 外周蛋白靠离子键或其它较弱的键与膜表面的蛋白质分子或脂分子的亲水部分结合,因 此只要改变溶液的离子强度甚至提高温度就可以从膜上分离下来,有时很难区分整合蛋白和外 周蛋白,主要是因为一个蛋白质可以由多个亚基构成,有的亚基为跨膜蛋白,有的则结合在膜 的外部。 脂锚定蛋白(lipid-anchored protein)可以分为两类,一类是糖磷脂酰肌醇 (glycophosphatidylinositol,,GPI)连接的蛋白,GPI位于细胞膜的外小叶,用磷脂酶C(能识 别含肌醇的磷脂)处理细胞,能释放出结合的蛋白。许多细胞表面的受体、酶、细胞粘附分子 和引起羊瘙痒病的PPC都是这类蛋白。另一类脂锚定蛋白与插入质膜内小叶的长碳氢链结 合,如三聚体GTP结合调节蛋白(trimeric GTP-binding regulatory protein)的a和Y亚基。 CYTOSOL 图4-11蛋白与膜的结合方式 ①、②整合蛋白;③、④脂锚定蛋白;⑤、⑥外周蛋白
图 4-10 Triton-X100 整合蛋白的跨膜结构域可以是 1 至多个疏水的 α 螺旋,形成亲水通道的整合蛋白跨膜区 域有两种组成形式,一是由多个两性 α 螺旋组成亲水通道;而是由两性 β 折叠组成亲水通道 (图 4-11)。 外周蛋白靠离子键或其它较弱的键与膜表面的蛋白质分子或脂分子的亲水部分结合,因 此只要改变溶液的离子强度甚至提高温度就可以从膜上分离下来,有时很难区分整合蛋白和外 周蛋白,主要是因为一个蛋白质可以由多个亚基构成,有的亚基为跨膜蛋白,有的则结合在膜 的外部。 脂锚定蛋白(lipid-anchored protein)可以分为两类,一类是糖磷脂酰肌醇 (glycophosphatidylinositol,GPI)连接的蛋白,GPI 位于细胞膜的外小叶,用磷脂酶 C(能识 别含肌醇的磷脂)处理细胞,能释放出结合的蛋白。许多细胞表面的受体、酶、细胞粘附分子 和引起羊瘙痒病的 PrPC 都是这类蛋白。另一类脂锚定蛋白与插入质膜内小叶的长碳氢链结 合,如三聚体 GTP 结合调节蛋白(trimeric GTP-binding regulatory protein)的 α 和 γ 亚基。 图 4-11 蛋白与膜的结合方式 ① 、②整合蛋白;③、④脂锚定蛋白;⑤、⑥外周蛋白

第二节质膜的结构 一、质膜结构的研究历史 1.E.Overton1895发现凡是溶于脂肪的物质很容易透过植物的细胞膜,而不溶于脂肪的 物质不易透过细胞膜,因此推测细胞膜由连续的脂类物质组成。 Hydrophilicmolecule Hydrophobic molecule 图4-12水溶性物质难以通过质膜 2.E.Gorter&F.Grendel1925用有机溶剂提取了人类红细胞质膜的脂类成分,将其铺 展在水面,测出膜脂展开的面积二倍于细胞表面积,因而推测细胞膜由双层脂分子组成。 3.J.Danielli&H.Davson1935发现质膜的表面张力比油-水界面的张力低得多,推测 膜中含有蛋白质,从而提出了”蛋白质-脂类-蛋白质”的三明治模型。认为质膜由双层脂类分子 及其内外表面附着的蛋白质构成的。1959年在上述基础上提出了修正模型,认为膜上还具有 贯穿脂双层的蛋白质通道,供亲水物质通过。 4.J.D.Robertson1959用超薄切片技术获得了清晰的细胞膜照片,显示暗-明-暗三层结 构(图4-13),厚约7.5nm。这就是所谓的“单位膜"模型。它由厚约3.5nm的双层脂分子和内外 表面各厚约2m的蛋白质构成。单位膜模型的不足之处在于把膜的动态结构描写成静止的不 变的
第二节 质膜的结构 一、质膜结构的研究历史 1. E. Overton 1895 发现凡是溶于脂肪的物质很容易透过植物的细胞膜,而不溶于脂肪的 物质不易透过细胞膜,因此推测细胞膜由连续的脂类物质组成。 图 4-12 水溶性物质难以通过质膜 2. E. Gorter & F. Grendel 1925 用有机溶剂提取了人类红细胞质膜的脂类成分,将其铺 展在水面,测出膜脂展开的面积二倍于细胞表面积,因而推测细胞膜由双层脂分子组成。 3. J. Danielli & H. Davson 1935 发现质膜的表面张力比油-水界面的张力低得多,推测 膜中含有蛋白质,从而提出了”蛋白质-脂类-蛋白质”的三明治模型。认为质膜由双层脂类分子 及其内外表面附着的蛋白质构成的。1959 年在上述基础上提出了修正模型,认为膜上还具有 贯穿脂双层的蛋白质通道,供亲水物质通过。 4. J. D. Robertson 1959 用超薄切片技术获得了清晰的细胞膜照片,显示暗-明-暗三层结 构(图 4-13),厚约 7.5nm。这就是所谓的“单位膜”模型。它由厚约 3.5nm 的双层脂分子和内外 表面各厚约 2nm 的蛋白质构成。单位膜模型的不足之处在于把膜的动态结构描写成静止的不 变的