
6 ③设计了细菌染色技术 (2)证实疾病的病原菌学说,提出了柯赫准则。 ①某一种微生物,当被怀疑是病原体时,它一定伴随着病害而存在; ②必须能自原寄主分离出这种微生物,并培养成为纯培养; ③用已纯化的纯培养微生物,人工接种寄主,必须能诱发与原来病害相同病害; ④必须自人工接种发病的寄主内,能重新分离出同一病原微生物并培养成纯培养。 其他人: Serge Winogradsky,1856~1953,发现微生物的自养生活; Beijerinck M. W.,1851~1931,发现了非共生固氮菌; Joseph Lister,1864 年提出无菌外科操作技术; Elie Metchnikoff 发现白细胞的吞噬作用; Ivanovsky 发现烟草花叶病毒; P. Ehrlich 现代化疗的开始。 3、现代微生物学发展—分子生物学阶段 (1)现代发酵工业的形成:1941,Florey & Chain 将青霉素投入生产,是通气培养微生物的开端,将微生物学与工程学结合。 (2)微生物代谢作用研究; 1944,Avery 肺炎球菌转化实验,确定 DNA 是遗传物质,标志着分子生物学的形成。 1953,Watson & Crick 提出 DNA 双螺旋结构以及半保留复制假说。 (3)分子生物学阶段 20 世纪 70 年代,基因工程的发展,工程菌的构建更促进了微生物学的发展。 微生物学推动生命科学的发展,促进许多重大理论问题的突破,对生命科学研究技术的贡献, 与"人类基因组计划"。 4、微生物学的应用前景 继续采用微生物作为生命科学的研究材料。 微生物生产与动植物生产并列为生物产业的三大支柱。 在工业中许多产品利用微生物来生产,如各种生物活性物质(抗生素等)、化工原料(酒精等)。 微生物在农业生产中也有着多方面的作用。 微生物在食品加工中有广泛用途,发酵食品和许多调味品都离不开微生物。 微生物是消除污染、净化环境的重要手段。 在新兴的生物技术产业中,微生物的作用更是不可替代。作为基因工程的外源 DNA 载体,不是 微生物本身(如噬菌体),就是微生物细胞中的质粒;被用作切割与拼接基因的工具酶,绝大多数来自 各种微生物。由于微生物生长繁殖快、培养条件较简易,当今大量的基因工程产品主要是以微生物作 为受体而进行生产,尤其是大肠杆菌、枯草芽胞杆菌和酿酒醉母。借助微生物发酵法,人们已能生产 外源蛋白质药物(如人胰岛素和干扰素等)。尽管基因工程所采用的外源基因可以来自动植物,但由于 微生物生理代谢类型的多样性,它们是最丰富的外源基因供体。 与高等动植物相比,已知微生物种类只是估计存在数量的很小一部分。哺乳动物和鸟类的物种几 乎全部为人们所掌握,被子植物已知种类达 93%,但细菌已知种数仅为估计数的 12%,真菌为 5%, 病毒为 4%(Bull,1992)。目前研究的也只是已知种类的很少一部分。根据 SCI(science citation index) 资料,1991—1997 发表的微生物学文献大量集中在 8 个属,尤其是埃希氏杆菌,其中大肠杆菌又占 主要部分(Galvez 等,1998)。可以想像,既然对少数已知微生物的研究就已为人类作出了重要贡献, 通过对多样性微生物的开发必然会为社会带来巨大利益。微生物学事业方兴未艾。 微生物基因组学研究将全面展开,以微生物之间、微生物与其他生物、微生物与环境的相互作用 为主要内容的微生物生态学、环境微生物学、细胞微生物学将基因组信息在基础上获得长足发展
6 ③设计了细菌染色技术 (2)证实疾病的病原菌学说,提出了柯赫准则。 ①某一种微生物,当被怀疑是病原体时,它一定伴随着病害而存在; ②必须能自原寄主分离出这种微生物,并培养成为纯培养; ③用已纯化的纯培养微生物,人工接种寄主,必须能诱发与原来病害相同病害; ④必须自人工接种发病的寄主内,能重新分离出同一病原微生物并培养成纯培养。 其他人: Serge Winogradsky,1856~1953,发现微生物的自养生活; Beijerinck M. W.,1851~1931,发现了非共生固氮菌; Joseph Lister,1864 年提出无菌外科操作技术; Elie Metchnikoff 发现白细胞的吞噬作用; Ivanovsky 发现烟草花叶病毒; P. Ehrlich 现代化疗的开始。 3、现代微生物学发展—分子生物学阶段 (1)现代发酵工业的形成:1941,Florey & Chain 将青霉素投入生产,是通气培养微生物的开端,将微生物学与工程学结合。 (2)微生物代谢作用研究; 1944,Avery 肺炎球菌转化实验,确定 DNA 是遗传物质,标志着分子生物学的形成。 1953,Watson & Crick 提出 DNA 双螺旋结构以及半保留复制假说。 (3)分子生物学阶段 20 世纪 70 年代,基因工程的发展,工程菌的构建更促进了微生物学的发展。 微生物学推动生命科学的发展,促进许多重大理论问题的突破,对生命科学研究技术的贡献, 与"人类基因组计划"。 4、微生物学的应用前景 继续采用微生物作为生命科学的研究材料。 微生物生产与动植物生产并列为生物产业的三大支柱。 在工业中许多产品利用微生物来生产,如各种生物活性物质(抗生素等)、化工原料(酒精等)。 微生物在农业生产中也有着多方面的作用。 微生物在食品加工中有广泛用途,发酵食品和许多调味品都离不开微生物。 微生物是消除污染、净化环境的重要手段。 在新兴的生物技术产业中,微生物的作用更是不可替代。作为基因工程的外源 DNA 载体,不是 微生物本身(如噬菌体),就是微生物细胞中的质粒;被用作切割与拼接基因的工具酶,绝大多数来自 各种微生物。由于微生物生长繁殖快、培养条件较简易,当今大量的基因工程产品主要是以微生物作 为受体而进行生产,尤其是大肠杆菌、枯草芽胞杆菌和酿酒醉母。借助微生物发酵法,人们已能生产 外源蛋白质药物(如人胰岛素和干扰素等)。尽管基因工程所采用的外源基因可以来自动植物,但由于 微生物生理代谢类型的多样性,它们是最丰富的外源基因供体。 与高等动植物相比,已知微生物种类只是估计存在数量的很小一部分。哺乳动物和鸟类的物种几 乎全部为人们所掌握,被子植物已知种类达 93%,但细菌已知种数仅为估计数的 12%,真菌为 5%, 病毒为 4%(Bull,1992)。目前研究的也只是已知种类的很少一部分。根据 SCI(science citation index) 资料,1991—1997 发表的微生物学文献大量集中在 8 个属,尤其是埃希氏杆菌,其中大肠杆菌又占 主要部分(Galvez 等,1998)。可以想像,既然对少数已知微生物的研究就已为人类作出了重要贡献, 通过对多样性微生物的开发必然会为社会带来巨大利益。微生物学事业方兴未艾。 微生物基因组学研究将全面展开,以微生物之间、微生物与其他生物、微生物与环境的相互作用 为主要内容的微生物生态学、环境微生物学、细胞微生物学将基因组信息在基础上获得长足发展

7 第二章 原核生物的形态、构造和功能 [教学目标]通过本章的教学,使学生掌握微生物的种类和特征、掌握以细菌为代表原核微生物的 形态结构、化学组成和繁殖特征等。 [教学的重点和难点]细菌的基本形态、构造和特征;原核微生物之间的区别;革兰氏染色;细菌 的群体形态特征。 [教学方法和手段]主要以讲授为主,应用多媒体课件进行形象生动的课堂教学。设计微生物学形 态学综合实验,从菌落形态、染色方法和显微镜观察等多方面进行实验教学。 [教学内容] 非细胞型(acellular microorganism):病毒、亚病毒 细胞型:原核微生物(prokaryotes):细菌、放线菌等。特点是无明显核,也无核膜、核仁。 原核生物即广义的细菌,指一大类细胞核无核膜包裹,只存在称作核区(nuclear region)的裸露 DNA 的原始单个细胞生物,包括真细菌(eubacteria)和古细菌(archaea)两大类群。 真核微生物(eukaryotic microorganism):酵母菌、霉菌。特点是有明显核,有核膜、核仁。 本章主要介绍原核生物的六种类型:细菌、放线菌、蓝细菌、支原体、立克次氏体和衣原体。 表 2.1 原核生物和真核生物遗传的和细胞组装上的主要差别 原核生物 真核生物 遗传物质和复制的组装 DNA 在细胞质中游离 DNA 在膜包围的核中,只有一个核仁 只有一个染色体 多于一个染色体,每个染色体是双拷贝(双倍体) DNA 与类组蛋白连系 DNA 与组蛋白连系 含有染色体外的遗传物质,称为质粒 只在酵母中发现质粒 在 mRNA 中没有发现内含子 所有基因中都发现内含子 细胞分裂以二等分裂方式,只有无性繁殖 细胞分裂为有丝分裂 遗传信息传递可通过接合、转导、转化发生 遗传信息交换发生在有性繁殖过程,减数分裂导致产生单倍体 细胞(配子),它们能融合。 细胞的组装 质膜含有 hopanoids、脂多糖和磷壁酸 质膜含有固醇 能量代谢与细胞质膜连系 多数情况在线粒体中发生 光合作用与细胞质中膜系统和泡囊连系 藻类和植物细胞中存在叶绿体 蛋白质合成和寻靶作用与内膜、粗糙内质网膜和高尔基体相连系 有膜的泡囊如溶酶体和过氧化物酶体有微管骨架存在 由一根蛋白鞭毛丝构成鞭毛 鞭毛有 9+2 微管排列的复杂结构 核糖体——70S 核糖体——80S(线粒体和叶绿体的核糖体是 70S) 肽聚糖的细胞壁(只有真细菌有,古细菌中是不同的多 聚体) 多糖的细胞壁,一般或者是纤维素或者是几丁质 原核细胞和真核细胞的区别 原核生物和真核生物 原核生物和真核生物细胞之间有许多差别。真核生物的主要特征是有细胞核和如线粒体、叶绿体 的细胞器及复杂的内膜系统。病毒属于非细胞类,细菌属于原核生物,所有其他微生物属于真核生物。 原核细胞和真核细胞的区别 核、核膜、染色体 原核生物细胞没有核膜,有一个明显的核区,这个核区上集中了它的主要遗传物质,由一条与类
7 第二章 原核生物的形态、构造和功能 [教学目标]通过本章的教学,使学生掌握微生物的种类和特征、掌握以细菌为代表原核微生物的 形态结构、化学组成和繁殖特征等。 [教学的重点和难点]细菌的基本形态、构造和特征;原核微生物之间的区别;革兰氏染色;细菌 的群体形态特征。 [教学方法和手段]主要以讲授为主,应用多媒体课件进行形象生动的课堂教学。设计微生物学形 态学综合实验,从菌落形态、染色方法和显微镜观察等多方面进行实验教学。 [教学内容] 非细胞型(acellular microorganism):病毒、亚病毒 细胞型:原核微生物(prokaryotes):细菌、放线菌等。特点是无明显核,也无核膜、核仁。 原核生物即广义的细菌,指一大类细胞核无核膜包裹,只存在称作核区(nuclear region)的裸露 DNA 的原始单个细胞生物,包括真细菌(eubacteria)和古细菌(archaea)两大类群。 真核微生物(eukaryotic microorganism):酵母菌、霉菌。特点是有明显核,有核膜、核仁。 本章主要介绍原核生物的六种类型:细菌、放线菌、蓝细菌、支原体、立克次氏体和衣原体。 表 2.1 原核生物和真核生物遗传的和细胞组装上的主要差别 原核生物 真核生物 遗传物质和复制的组装 DNA 在细胞质中游离 DNA 在膜包围的核中,只有一个核仁 只有一个染色体 多于一个染色体,每个染色体是双拷贝(双倍体) DNA 与类组蛋白连系 DNA 与组蛋白连系 含有染色体外的遗传物质,称为质粒 只在酵母中发现质粒 在 mRNA 中没有发现内含子 所有基因中都发现内含子 细胞分裂以二等分裂方式,只有无性繁殖 细胞分裂为有丝分裂 遗传信息传递可通过接合、转导、转化发生 遗传信息交换发生在有性繁殖过程,减数分裂导致产生单倍体 细胞(配子),它们能融合。 细胞的组装 质膜含有 hopanoids、脂多糖和磷壁酸 质膜含有固醇 能量代谢与细胞质膜连系 多数情况在线粒体中发生 光合作用与细胞质中膜系统和泡囊连系 藻类和植物细胞中存在叶绿体 蛋白质合成和寻靶作用与内膜、粗糙内质网膜和高尔基体相连系 有膜的泡囊如溶酶体和过氧化物酶体有微管骨架存在 由一根蛋白鞭毛丝构成鞭毛 鞭毛有 9+2 微管排列的复杂结构 核糖体——70S 核糖体——80S(线粒体和叶绿体的核糖体是 70S) 肽聚糖的细胞壁(只有真细菌有,古细菌中是不同的多 聚体) 多糖的细胞壁,一般或者是纤维素或者是几丁质 原核细胞和真核细胞的区别 原核生物和真核生物 原核生物和真核生物细胞之间有许多差别。真核生物的主要特征是有细胞核和如线粒体、叶绿体 的细胞器及复杂的内膜系统。病毒属于非细胞类,细菌属于原核生物,所有其他微生物属于真核生物。 原核细胞和真核细胞的区别 核、核膜、染色体 原核生物细胞没有核膜,有一个明显的核区,这个核区上集中了它的主要遗传物质,由一条与类

8 组蛋白相联系的双链 DNA 构成的染色体组成。 真核生物细胞则是由一条或一条以上的双链 DNA 与组蛋白等结合成的染色体,并由核膜包围。 代谢场所 原核细胞没有独立的内膜系统,与代谢有关的酶如呼吸酶合成酶等位于细胞膜上,因此它的能量 代谢在质膜上进行。 真核细胞不仅有独立的内膜系统,还有细胞骨架,呼吸酶在线粒体中,有专用的细胞器来完成各 项生理功能,如线粒体、叶绿体。 核糖体的大小和分布 原核细胞的核糖体大小为 70S,常以游离状态或多聚体状态分布于细胞质中。 真核细胞的核糖体大小为 80S,可以游离状态存在于细胞结合于内质网上。线粒体和叶绿体内有 各自在结构上特殊的核糖体。 §1 细菌 细菌(bacteria)是一类细胞细短(直径约 0.5μm,长度约 0.5~5μm)、结构简单,胞壁坚韧, 多以二分裂方式繁殖和水生性较强的原核生物。1000 倍以上显微镜才能看到其形状。 一、细菌的形态构造及其功能 (一)形态与染色 1、基本外形:球状——球菌;杆状——杆菌;螺旋状——螺旋菌。 (1)球菌(Coccus): 球形或近球形,根据空间排列方式不同又分为单、双、链、四联、八叠、葡萄球菌。不同的排列 方式是由于细胞分裂方向及分裂后情况不同造成的。细胞呈球状或椭圆形。根据这些细胞分裂产生的 新细胞所保持的一定空间排列方式有以下几种情形:见图 2-1 单球菌——尿素微球菌(图 2-1-1) 双球菌——肺炎双球菌(图 2-1-2) 链球菌——溶血链球菌(图 2-1-3) 四联球菌——四联微球菌(图 2-1-4) 八叠球菌——尿素八叠球菌(图 2-1-5) 葡萄球菌——金黄色 葡萄球菌(图 2-1-6) (2)杆菌 Bacillus (Bacterium) :杆状 或圆柱形,径长比不同,短粗或细长。是细菌中种类最多的。 杆菌细胞呈杆状或圆柱形。图 2.1 中 B 的 7 为长杆菌和短杆菌,8 为枯草芽孢杆菌,9 为溶纤维梭菌。 (3)螺旋菌(Spirillum):细胞呈弯曲杆状的细菌统称为螺旋菌。是细胞呈弯曲杆状细菌统称, 一般分散存在。根据其长度、螺旋数目和螺距等差别,分为弧菌 Vibrio(菌体只有一个弯曲,形似 C 字)和螺旋菌(螺旋状,超过 1 圈)。 与螺旋体 Spirochaeta 区别:无鞭毛。 弧菌 偏端单生鞭毛或丛生鞭毛(图 2-1-10) 螺旋菌 两端都有鞭毛(图 2-1-11) 细菌形态不是一成不变的,受环境条件影响(如温度、培养基浓度及组成、菌龄等)。 异常形态:一般,幼龄,生长条件适宜,形状正常、整齐。老龄,不正常,异常形态。 畸形:由于理化因素刺激,阻碍细胞发育引起。 衰颓形:由于培养时间长,细胞衰老,营养缺乏,或排泄物积累过多引起。 2、细菌染色法 由于细菌细胞既小又透明,故一般先要经过染色才能作显微镜观察
8 组蛋白相联系的双链 DNA 构成的染色体组成。 真核生物细胞则是由一条或一条以上的双链 DNA 与组蛋白等结合成的染色体,并由核膜包围。 代谢场所 原核细胞没有独立的内膜系统,与代谢有关的酶如呼吸酶合成酶等位于细胞膜上,因此它的能量 代谢在质膜上进行。 真核细胞不仅有独立的内膜系统,还有细胞骨架,呼吸酶在线粒体中,有专用的细胞器来完成各 项生理功能,如线粒体、叶绿体。 核糖体的大小和分布 原核细胞的核糖体大小为 70S,常以游离状态或多聚体状态分布于细胞质中。 真核细胞的核糖体大小为 80S,可以游离状态存在于细胞结合于内质网上。线粒体和叶绿体内有 各自在结构上特殊的核糖体。 §1 细菌 细菌(bacteria)是一类细胞细短(直径约 0.5μm,长度约 0.5~5μm)、结构简单,胞壁坚韧, 多以二分裂方式繁殖和水生性较强的原核生物。1000 倍以上显微镜才能看到其形状。 一、细菌的形态构造及其功能 (一)形态与染色 1、基本外形:球状——球菌;杆状——杆菌;螺旋状——螺旋菌。 (1)球菌(Coccus): 球形或近球形,根据空间排列方式不同又分为单、双、链、四联、八叠、葡萄球菌。不同的排列 方式是由于细胞分裂方向及分裂后情况不同造成的。细胞呈球状或椭圆形。根据这些细胞分裂产生的 新细胞所保持的一定空间排列方式有以下几种情形:见图 2-1 单球菌——尿素微球菌(图 2-1-1) 双球菌——肺炎双球菌(图 2-1-2) 链球菌——溶血链球菌(图 2-1-3) 四联球菌——四联微球菌(图 2-1-4) 八叠球菌——尿素八叠球菌(图 2-1-5) 葡萄球菌——金黄色 葡萄球菌(图 2-1-6) (2)杆菌 Bacillus (Bacterium) :杆状 或圆柱形,径长比不同,短粗或细长。是细菌中种类最多的。 杆菌细胞呈杆状或圆柱形。图 2.1 中 B 的 7 为长杆菌和短杆菌,8 为枯草芽孢杆菌,9 为溶纤维梭菌。 (3)螺旋菌(Spirillum):细胞呈弯曲杆状的细菌统称为螺旋菌。是细胞呈弯曲杆状细菌统称, 一般分散存在。根据其长度、螺旋数目和螺距等差别,分为弧菌 Vibrio(菌体只有一个弯曲,形似 C 字)和螺旋菌(螺旋状,超过 1 圈)。 与螺旋体 Spirochaeta 区别:无鞭毛。 弧菌 偏端单生鞭毛或丛生鞭毛(图 2-1-10) 螺旋菌 两端都有鞭毛(图 2-1-11) 细菌形态不是一成不变的,受环境条件影响(如温度、培养基浓度及组成、菌龄等)。 异常形态:一般,幼龄,生长条件适宜,形状正常、整齐。老龄,不正常,异常形态。 畸形:由于理化因素刺激,阻碍细胞发育引起。 衰颓形:由于培养时间长,细胞衰老,营养缺乏,或排泄物积累过多引起。 2、细菌染色法 由于细菌细胞既小又透明,故一般先要经过染色才能作显微镜观察
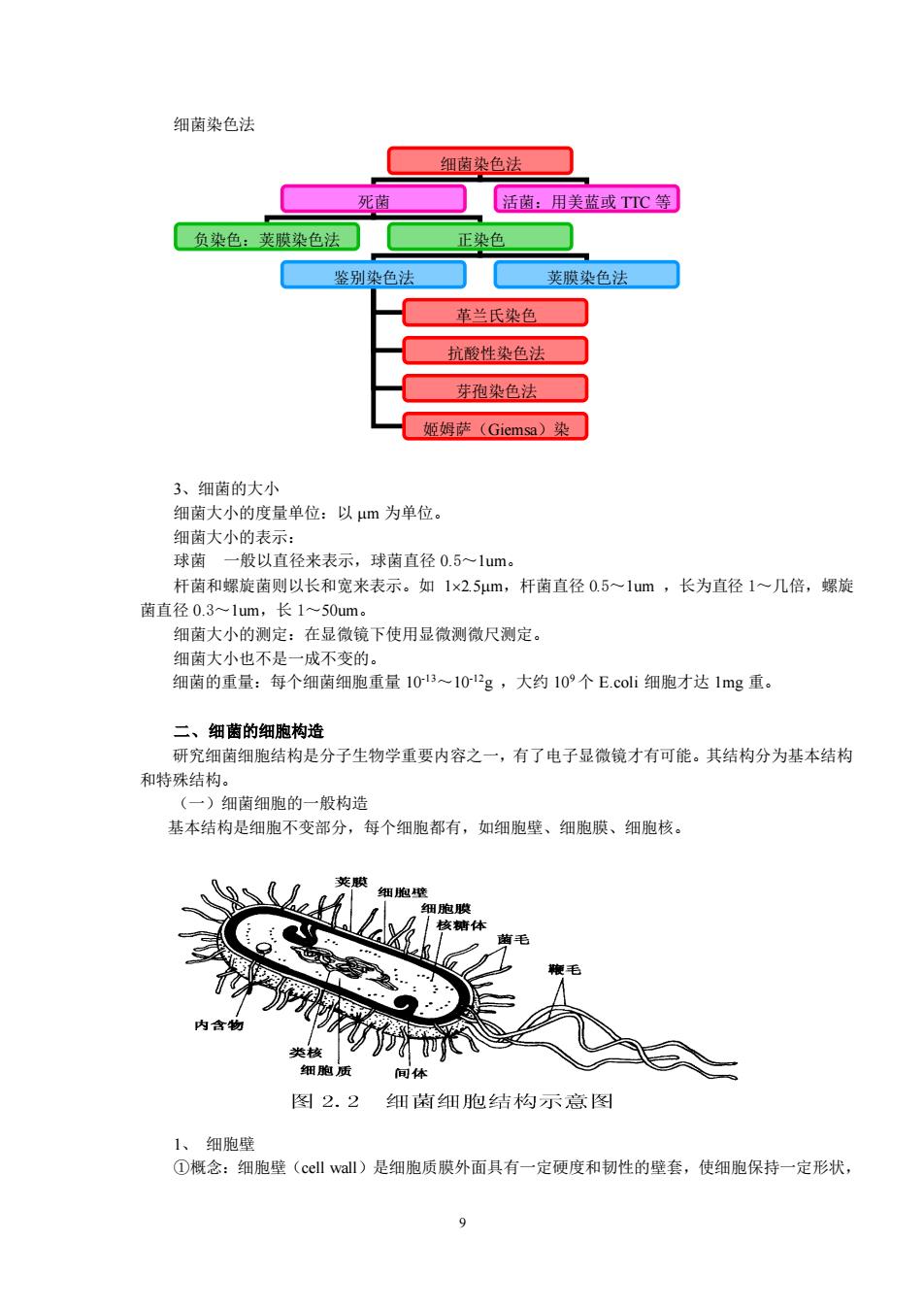
9 细菌染色法 3、细菌的大小 细菌大小的度量单位:以 m 为单位。 细菌大小的表示: 球菌 一般以直径来表示,球菌直径 0.5~1um。 杆菌和螺旋菌则以长和宽来表示。如 12.5m,杆菌直径 0.5~1um ,长为直径 1~几倍,螺旋 菌直径 0.3~1um,长 1~50um。 细菌大小的测定:在显微镜下使用显微测微尺测定。 细菌大小也不是一成不变的。 细菌的重量:每个细菌细胞重量 10-13~10-12g ,大约 109 个 E.coli 细胞才达 1mg 重。 二、细菌的细胞构造 研究细菌细胞结构是分子生物学重要内容之一,有了电子显微镜才有可能。其结构分为基本结构 和特殊结构。 (一)细菌细胞的一般构造 基本结构是细胞不变部分,每个细胞都有,如细胞壁、细胞膜、细胞核。 1、 细胞壁 ①概念:细胞壁(cell wall)是细胞质膜外面具有一定硬度和韧性的壁套,使细胞保持一定形状, 细菌染色法 死菌 活菌:用美蓝或 TTC 等 负染色:荚膜染色法 正染色 鉴别染色法 荚膜染色法 革兰氏染色 抗酸性染色法 芽孢染色法 姬姆萨(Giemsa)染 色法
9 细菌染色法 3、细菌的大小 细菌大小的度量单位:以 m 为单位。 细菌大小的表示: 球菌 一般以直径来表示,球菌直径 0.5~1um。 杆菌和螺旋菌则以长和宽来表示。如 12.5m,杆菌直径 0.5~1um ,长为直径 1~几倍,螺旋 菌直径 0.3~1um,长 1~50um。 细菌大小的测定:在显微镜下使用显微测微尺测定。 细菌大小也不是一成不变的。 细菌的重量:每个细菌细胞重量 10-13~10-12g ,大约 109 个 E.coli 细胞才达 1mg 重。 二、细菌的细胞构造 研究细菌细胞结构是分子生物学重要内容之一,有了电子显微镜才有可能。其结构分为基本结构 和特殊结构。 (一)细菌细胞的一般构造 基本结构是细胞不变部分,每个细胞都有,如细胞壁、细胞膜、细胞核。 1、 细胞壁 ①概念:细胞壁(cell wall)是细胞质膜外面具有一定硬度和韧性的壁套,使细胞保持一定形状, 细菌染色法 死菌 活菌:用美蓝或 TTC 等 负染色:荚膜染色法 正染色 鉴别染色法 荚膜染色法 革兰氏染色 抗酸性染色法 芽孢染色法 姬姆萨(Giemsa)染 色法

10 保障其在不同渗透压条件下生长,即使在不良环境中也能防止胞溶作用。 真细菌的细胞壁由肽聚糖构成,而古细菌细胞壁组成物质极为多样,从类似肽聚糖的物质、假肽 聚糖,到多糖、蛋白质和糖蛋白。 真细菌细胞壁由肤聚糖构成,肤聚糖是 N-乙酰氨基葡萄糖(NAG)和带有交替排列的 D-型或 L-型 氨基酸侧链的 N-乙酰胞壁酸(NAM)的多聚体。它是高度的交联的分子,使得细胞具有刚性、强度和 保护细胞抵抗渗透压的裂解。肽聚糖有许多独特的特性,如 D-型氨基酸,它可作为抗生素攻击肽聚 糖的靶目标(抗生素通过抑制或干扰肽聚糖合成而使细胞壁缺损)。革兰氏阳性细菌细胞还含有磷壁酸。 ②功能: 细菌细胞壁的生理功能有: 保护原生质体免受渗透压引起破裂的作用;维持细菌的细胞形态(可用溶菌酶处理不同形态的细 菌细胞壁后,菌体均呈现圆形得到证明);细胞壁是多孔结构的分子筛,阻挡某些分子进入和保留蛋 白质在间质(革兰氏阴性菌细胞壁和细胞质之间的区域);细胞壁为鞭毛提供支点,使鞭毛运动。 ③革兰氏染色 革兰氏染色根据 1884 年革兰姆·克里斯琴(Christian Gram)发明的染色反应,真细菌常常分成两 类。对染色步骤反应的差别是由于两类细菌的细胞外膜结构。革兰氏阳性细菌有单一的膜称作细胞膜 (或原生质膜),周围被厚的肽聚糖层包围(20~80nm)。革兰氏阴性细菌只有一薄层肽聚糖(1—3nm), 但是在肽聚糖层外边,仍有另一层的外膜,作为另外的屏障(图 2.3)。 革兰氏染色步骤如下:固定过的细胞用暗染色例如结晶紫染色,接着加碘液媒染,细菌 细胞壁内由于染色形成结晶紫与碘的复合物。随后加酒精从薄的细胞壁中洗出结晶紫与碘暗 染色的复合物,但是结晶紫—碘复合物不能从厚的细胞壁中洗出。最后,用较浅的石炭酸复 红复染。加石炭酸复红染色,使脱色的细胞呈粉红色,但在暗染色的细胞中没有看到粉红色, 仍保持第一次的染色结果。保持原来染色(厚的细胞壁)的细胞称作革兰氏阳性,在光学显微 镜下呈现蓝紫色。脱色的细胞(薄的细胞壁和外膜)称作革兰氏阴性,染成粉红色或淡紫色。 表 2.2 革兰氏染色程序和结果 步 骤 方 法 结 果 阳性(G+) 阴性(G-) 初 染 结晶紫 30s 紫 色 紫 色 媒染剂 碘液 30s 仍为紫色 仍为紫色 脱 色 95%乙醇 10—20s 保持紫色 脱去紫色 复 染 蕃红(或复红)30—60s 仍显紫色 红 色 ④化学组成与超微结构 a 革兰氏阳性细菌(Gram positive) 革兰氏阳性细菌细胞壁具有较厚(30~40nm)而致密的肽聚糖层,多达 20 层,占细胞壁的成分 60-90%,它同细胞膜的外层紧密相连(见图 2.4)。 有的革兰氏阳性细菌细胞壁中含有磷壁酸(teichoi-acid),也即胞壁质(murein)
10 保障其在不同渗透压条件下生长,即使在不良环境中也能防止胞溶作用。 真细菌的细胞壁由肽聚糖构成,而古细菌细胞壁组成物质极为多样,从类似肽聚糖的物质、假肽 聚糖,到多糖、蛋白质和糖蛋白。 真细菌细胞壁由肤聚糖构成,肤聚糖是 N-乙酰氨基葡萄糖(NAG)和带有交替排列的 D-型或 L-型 氨基酸侧链的 N-乙酰胞壁酸(NAM)的多聚体。它是高度的交联的分子,使得细胞具有刚性、强度和 保护细胞抵抗渗透压的裂解。肽聚糖有许多独特的特性,如 D-型氨基酸,它可作为抗生素攻击肽聚 糖的靶目标(抗生素通过抑制或干扰肽聚糖合成而使细胞壁缺损)。革兰氏阳性细菌细胞还含有磷壁酸。 ②功能: 细菌细胞壁的生理功能有: 保护原生质体免受渗透压引起破裂的作用;维持细菌的细胞形态(可用溶菌酶处理不同形态的细 菌细胞壁后,菌体均呈现圆形得到证明);细胞壁是多孔结构的分子筛,阻挡某些分子进入和保留蛋 白质在间质(革兰氏阴性菌细胞壁和细胞质之间的区域);细胞壁为鞭毛提供支点,使鞭毛运动。 ③革兰氏染色 革兰氏染色根据 1884 年革兰姆·克里斯琴(Christian Gram)发明的染色反应,真细菌常常分成两 类。对染色步骤反应的差别是由于两类细菌的细胞外膜结构。革兰氏阳性细菌有单一的膜称作细胞膜 (或原生质膜),周围被厚的肽聚糖层包围(20~80nm)。革兰氏阴性细菌只有一薄层肽聚糖(1—3nm), 但是在肽聚糖层外边,仍有另一层的外膜,作为另外的屏障(图 2.3)。 革兰氏染色步骤如下:固定过的细胞用暗染色例如结晶紫染色,接着加碘液媒染,细菌 细胞壁内由于染色形成结晶紫与碘的复合物。随后加酒精从薄的细胞壁中洗出结晶紫与碘暗 染色的复合物,但是结晶紫—碘复合物不能从厚的细胞壁中洗出。最后,用较浅的石炭酸复 红复染。加石炭酸复红染色,使脱色的细胞呈粉红色,但在暗染色的细胞中没有看到粉红色, 仍保持第一次的染色结果。保持原来染色(厚的细胞壁)的细胞称作革兰氏阳性,在光学显微 镜下呈现蓝紫色。脱色的细胞(薄的细胞壁和外膜)称作革兰氏阴性,染成粉红色或淡紫色。 表 2.2 革兰氏染色程序和结果 步 骤 方 法 结 果 阳性(G+) 阴性(G-) 初 染 结晶紫 30s 紫 色 紫 色 媒染剂 碘液 30s 仍为紫色 仍为紫色 脱 色 95%乙醇 10—20s 保持紫色 脱去紫色 复 染 蕃红(或复红)30—60s 仍显紫色 红 色 ④化学组成与超微结构 a 革兰氏阳性细菌(Gram positive) 革兰氏阳性细菌细胞壁具有较厚(30~40nm)而致密的肽聚糖层,多达 20 层,占细胞壁的成分 60-90%,它同细胞膜的外层紧密相连(见图 2.4)。 有的革兰氏阳性细菌细胞壁中含有磷壁酸(teichoi-acid),也即胞壁质(murein)