
上丙丽度脱搬形成的整艺基得生物与丙 3作为德幸酸装减麻的乙最盖转夏 质碳结合 轴南A HC-000 CS-LSH CHr C-S-CoA SH HC阳-TP FAD ”氧辛 氧化魔基德率膜,使氧化型成 再生 NAD NADH+H' 三二氢魔事骤脱氧瑞使还球型黄素道 白量颜氧化 图4.4丙酮酸脱氢酶复合物的作用机制 丙弱酸十CoASH十NADt丙丽酸服复台林乙酰COA十CO,十NADH十H 2.三羧酸循环(tricarboxylic acid cycle) 乙酰CoA进入由一连串反应构成的循环体系,被氧化生成H2O和CO2。由于这个循环反应开始于乙酰CoA与草酰乙酸(oxaloacetate)缩合生成的 含有三个羧基的柠檬酸,因此称之为三羧酸循环或柠檬酸循环(citric acid cycle)。其详细过程如下: (1)乙酰CoA进入三羧酸循环 乙酰C0A具有硫酯键,乙酰基有足够能量与草酰乙酸的羧基进行醛醇型缩合。首先从CH3CO基上除去一个H+,生成的阴离子对草酰乙酸的羰 基碳进行亲核攻击,生成柠檬酰C0A中间体,然后高能硫酯键水解放出游离的柠檬酸,使反应不可逆地向右进行。该反应由柠檬酸合成酶 (citrate synthetase)催化,是很强的放能反应. 乙酰CoA+草酰乙酸柠谦联合皮鹰柠檬酸+CoASH 由草酰乙酸和乙酰C0A合成柠檬酸是三羧酸循环的重要调节点,柠檬酸合成酶是一个变构酶,ATP是柠檬酸合成酶的变构抑制剂,此外,α-酮 戊二酸、NADH能变构抑制其活性,长链脂酰CoA也可抑制它的活性,AMP可对抗ATP的抑制而起激活作用。 (2)异柠檬酸形成 柠檬酸的叔醇基不易氧化,转变成异柠檬酸而使叔醇变成仲醇,就易于氧化,此反应由顺乌头酸酶催化,为一可逆反应。 柠檬酸一顺飞头酸群异柠檬酸 (3)第一次氧化脱酸 在异柠檬酸脱氢酶作用下,异柠檬酸的仲醇氧化成羰基,生成草酰琥珀酸(oxalosuccinate)的中间产物,后者在同一酶表面,快速脱羧生成a-酮 戊二酸(a?ketoglutarate)、NADH和CO2,此反应为B-氧化脱羧,此酶需要Mn2+作为激活剂 异柠膝酸+NAD+界柠速酸氢恶e酮成二酸+CO,十NADH+H时 此反应是不可逆的,是三羧酸循环中的限速步骤,ADP是异柠檬酸脱氢酶的激活剂,而ATP,NADH是此酶的抑制剂. (4)第二次氧化脱羧 在α-酮戊二酸脱氢酶系作用下,α-酮戊二酸氧化脱羧生成琥珀酰CoA、NADH+H+和CO2,反应过程完全类似于丙酮酸脱氢酶系催化的氧化脱 羧,属于α?氧化脱羧,氧化产生的能量中一部分储存于琥珀酰CoA的高能硫酯键中。 α-酮戊二酸脱氢酶系也由三个酶(α-酮戊二酸脱羧酶、硫辛酸琥珀酰基转移酶、二氢硫辛酸脱氢酶)和五个辅酶(TPP、硫辛酸、HSCoA、 NAD+、FAD)组成 。-南皮二骏+CoASH+NAD~我三酸氢器我珀酰COA十NADH+H+CO, 此反应也是不可逆的。a-酮戊二酸脱氢酶复合体受ATP、GTP、NAPH和琥珀酰CoA抑制,但其不受磷酸化/去磷酸化的调控。 (5)底物磷酸化生成ATP 在琥珀酸硫激酶(succinatethiokinase)的作用下,琥珀酰CoA的硫酯键水解,释放的自由能用于合成GTP,在细菌和高等生物可直接生成ATP 在哺乳动物中,先生成GTP,再生成ATP,此时,琥珀酰CoA生成琥珀酸和辅酶A。 號珀酰CoA十GDP,乳珀眼合缆珀酸十HSCoA-十GTP (6)琥珀酸脱氢 琥珀酸脱氢酶(succinatedehydrogenase)催化琥珀酸氧化成为延胡索酸。该酶结合在线粒体内膜上,而其他三羧酸循环的酶则都是存在线粒体基 质中的,这酶含有铁硫中心和共价结合的FAD,来自琥珀酸的电子通过FAD和铁硫中心,然后进入电子传递链到O2,丙二酸是琥珀酸的类似
图4-4 丙酮酸脱氢酶复合物的作用机制 2.三羧酸循环(tricarboxylic acid cycle) 乙酰CoA进入由一连串反应构成的循环体系,被氧化生成H2O和CO2。由于这个循环反应开始于乙酰CoA与草酰乙酸(oxaloacetate)缩合生成的 含有三个羧基的柠檬酸,因此称之为三羧酸循环或柠檬酸循环(citric acid cycle)。其详细过程如下: (1)乙酰CoA进入三羧酸循环 乙酰CoA具有硫酯键,乙酰基有足够能量与草酰乙酸的羧基进行醛醇型缩合。首先从CH3CO基上除去一个H+,生成的阴离子对草酰乙酸的羰 基碳进行亲核攻击,生成柠檬酰CoA中间体,然后高能硫酯键水解放出游离的柠檬酸,使反应不可逆地向右进行。该反应由柠檬酸合成酶 (citrate synthetase)催化,是很强的放能反应。 由草酰乙酸和乙酰CoA合成柠檬酸是三羧酸循环的重要调节点,柠檬酸合成酶是一个变构酶,ATP是柠檬酸合成酶的变构抑制剂,此外,α-酮 戊二酸、NADH能变构抑制其活性,长链脂酰CoA也可抑制它的活性,AMP可对抗ATP的抑制而起激活作用。 (2)异柠檬酸形成 柠檬酸的叔醇基不易氧化,转变成异柠檬酸而使叔醇变成仲醇,就易于氧化,此反应由顺乌头酸酶催化,为一可逆反应。 (3)第一次氧化脱酸 在异柠檬酸脱氢酶作用下,异柠檬酸的仲醇氧化成羰基,生成草酰琥珀酸(oxalosuccinate)的中间产物,后者在同一酶表面,快速脱羧生成α-酮 戊二酸(α?ketoglutarate)、NADH和CO2,此反应为β-氧化脱羧,此酶需要Mn2+作为激活剂。 此反应是不可逆的,是三羧酸循环中的限速步骤,ADP是异柠檬酸脱氢酶的激活剂,而ATP,NADH是此酶的抑制剂。 (4)第二次氧化脱羧 在α-酮戊二酸脱氢酶系作用下,α-酮戊二酸氧化脱羧生成琥珀酰CoA、NADH+H+和CO2,反应过程完全类似于丙酮酸脱氢酶系催化的氧化脱 羧,属于α?氧化脱羧,氧化产生的能量中一部分储存于琥珀酰CoA的高能硫酯键中。 α-酮戊二酸脱氢酶系也由三个酶(α-酮戊二酸脱羧酶、硫辛酸琥珀酰基转移酶、二氢硫辛酸脱氢酶)和五个辅酶(TPP、硫辛酸、HSCoA、 NAD+、FAD)组成。 此反应也是不可逆的。α-酮戊二酸脱氢酶复合体受ATP、GTP、NAPH和琥珀酰CoA抑制,但其不受磷酸化/去磷酸化的调控。 (5)底物磷酸化生成ATP 在琥珀酸硫激酶(succinatethiokinase)的作用下,琥珀酰CoA的硫酯键水解,释放的自由能用于合成GTP,在细菌和高等生物可直接生成ATP, 在哺乳动物中,先生成GTP,再生成ATP,此时,琥珀酰CoA生成琥珀酸和辅酶A。 (6)琥珀酸脱氢 琥珀酸脱氢酶(succinatedehydrogenase)催化琥珀酸氧化成为延胡索酸。该酶结合在线粒体内膜上,而其他三羧酸循环的酶则都是存在线粒体基 质中的,这酶含有铁硫中心和共价结合的FAD,来自琥珀酸的电子通过FAD和铁硫中心,然后进入电子传递链到O2,丙二酸是琥珀酸的类似
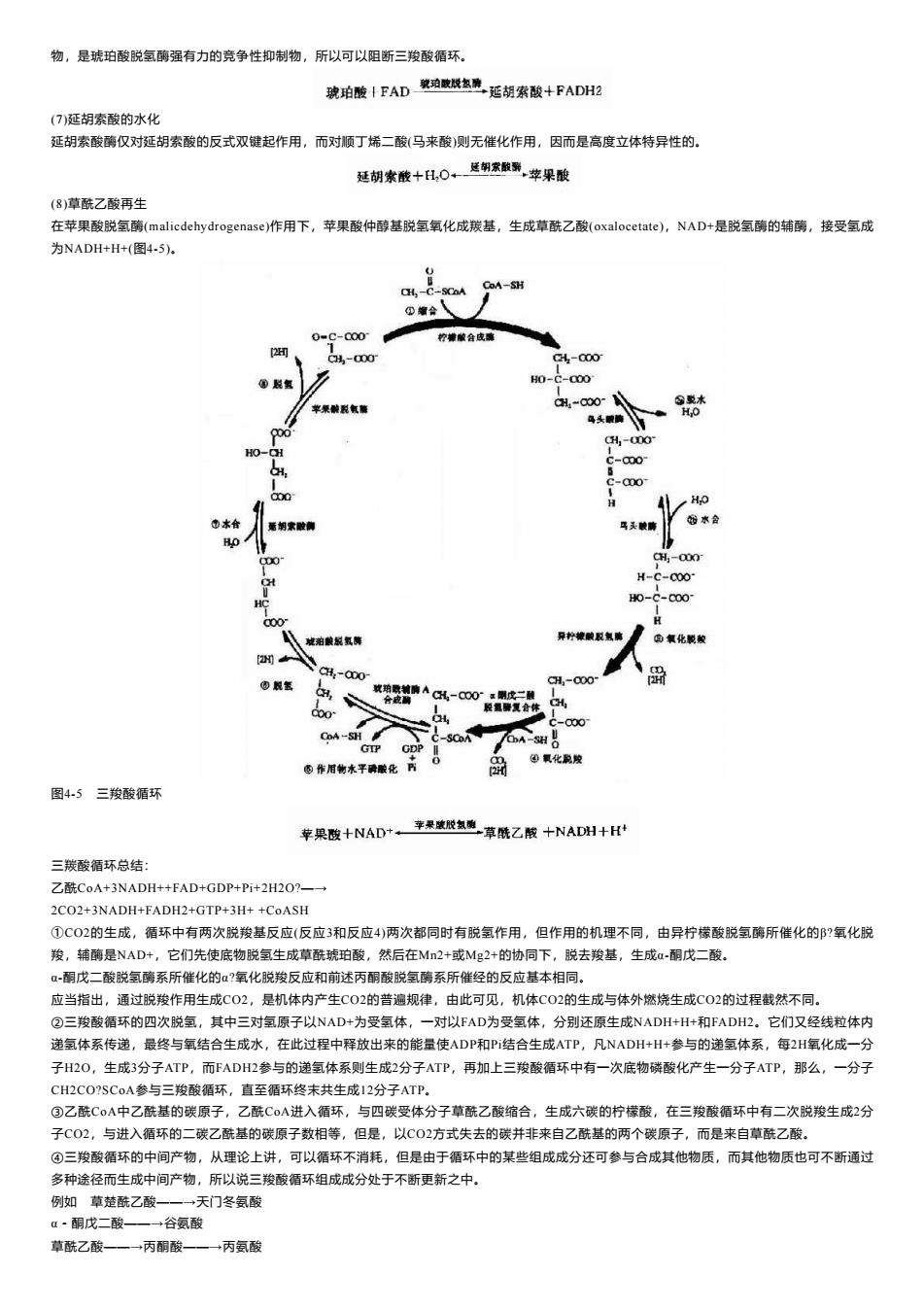
物,是琥珀酸脱氢酶强有力的竞争性抑制物,所以可以阻断三羧酸循环。 琥珀酸十FAD珀威氢麂延胡素酸+FADH2 (7)延胡索酸的水化 延胡索酸酶仅对延胡索酸的反式双键起作用,而对顺丁烯二酸(马来酸)则无催化作用,因而是高度立体特异性的。 延胡索酸十H,0←翻家服影苹果酸 (8)草酰乙酸再生 在苹果酸脱氢酶(malicdehydrogenase)作用下,苹果酸仲醇基脱氢氧化成羰基,生成草酰乙酸(oxalocetate),NAD+是脱氢酶的辅酶,接受氢成 为NADH+H+(图4-5). CH-C-SCoA CoA-SH ④缩合、 0=C-Q00 柠神酸合成囊 2 4-00 4-000 H0-c-a00 H-a00 ⊙乾水 苹聚酸税氧酶 鸟头眼喷 H,0 1-00 c-00 c-a00 1 H HO 合 马头酸商 水合 H,-000 H-C-C0O- H0-C-c00° Q00 装珀酸脱氧算 向氧化乾酸 山限氢 CH:-000 aH,-000 效珀歌轴情ACH-C00:明成二融 合应 经氢聘夏合休 CH, 00 c-000 CoA-S明 C-SCDA GTP GDP 0 ④氧化脱陵 西作用物水平聘酸化野 2可 图4-5三羧酸循环 幸果险十NAD+,率果威氢题草酰乙酸十NADH十H 三羰酸循环总结: 乙酰CoA+3NADH++FAD+GDP+Pi+2H2O?一→ 2C02+3NADH+FADH2+GTP+3H++CoASH ①C02的生成,循环中有两次脱羧基反应(反应3和反应4)两次都同时有脱氢作用,但作用的机理不同,由异柠檬酸脱氢酶所催化的?氧化脱 羧,辅酶是NAD+,它们先使底物脱氢生成草酰琥珀酸,然后在M2+或Mg2+的协同下,脱去羧基,生成a-酮戊二酸。 α-酮戊二酸脱氢酶系所催化的α?氧化脱羧反应和前述丙酮酸脱氢酶系所催经的反应基本相同。 应当指出,通过脱羧作用生成CO2,是机体内产生C02的普遍规律,由此可见,机体CO2的生成与体外燃烧生成CO2的过程截然不同。 ②三羧酸循环的四次脱氢,其中三对氢原子以NAD+为受氢体,一对以FAD为受氢体,分别还原生成NADH+H+和FADH2。它们又经线粒体内 递氢体系传递,最终与氧结合生成水,在此过程中释放出来的能量使ADP和P结合生成ATP,凡NADH+H+参与的递氢体系,每2H氧化成一分 子H2O,生成3分子ATP,而FADH2参与的递氢体系则生成2分子ATP,再加上三羧酸循环中有一次底物磷酸化产生一分子ATP,那么,一分子 CH2CO?SCoA参与三羧酸循环,直至循环终末共生成12分子ATP, ③乙酰CoA中乙酰基的碳原子,乙酰C0A进入循环,与四碳受体分子草酰乙酸缩合,生成六碳的柠檬酸,在三羧酸循环中有二次脱羧生成2分 子C02,与进入循环的二碳乙酰基的碳原子数相等,但是,以C02方式失去的碳并非来自乙酰基的两个碳原子,而是来自草酰乙酸。 ④三羧酸循环的中间产物,从理论上讲,可以循环不消耗,但是由于循环中的某些组成成分还可参与合成其他物质,而其他物质也可不断通过 多种途径而生成中间产物,所以说三羧酸循环组成成分处于不断更新之中。 例如草楚酰乙酸一一→天门冬氨酸 α·酮戊二酸一一→+谷氨酸 草酰乙酸一一→丙酮酸一一→丙氨酸
物,是琥珀酸脱氢酶强有力的竞争性抑制物,所以可以阻断三羧酸循环。 (7)延胡索酸的水化 延胡索酸酶仅对延胡索酸的反式双键起作用,而对顺丁烯二酸(马来酸)则无催化作用,因而是高度立体特异性的。 (8)草酰乙酸再生 在苹果酸脱氢酶(malicdehydrogenase)作用下,苹果酸仲醇基脱氢氧化成羰基,生成草酰乙酸(oxalocetate),NAD+是脱氢酶的辅酶,接受氢成 为NADH+H+(图4-5)。 图4-5 三羧酸循环 三羰酸循环总结: 乙酰CoA+3NADH++FAD+GDP+Pi+2H2O?—→ 2CO2+3NADH+FADH2+GTP+3H+ +CoASH ①CO2的生成,循环中有两次脱羧基反应(反应3和反应4)两次都同时有脱氢作用,但作用的机理不同,由异柠檬酸脱氢酶所催化的β?氧化脱 羧,辅酶是NAD+,它们先使底物脱氢生成草酰琥珀酸,然后在Mn2+或Mg2+的协同下,脱去羧基,生成α-酮戊二酸。 α-酮戊二酸脱氢酶系所催化的α?氧化脱羧反应和前述丙酮酸脱氢酶系所催经的反应基本相同。 应当指出,通过脱羧作用生成CO2,是机体内产生CO2的普遍规律,由此可见,机体CO2的生成与体外燃烧生成CO2的过程截然不同。 ②三羧酸循环的四次脱氢,其中三对氢原子以NAD+为受氢体,一对以FAD为受氢体,分别还原生成NADH+H+和FADH2。它们又经线粒体内 递氢体系传递,最终与氧结合生成水,在此过程中释放出来的能量使ADP和Pi结合生成ATP,凡NADH+H+参与的递氢体系,每2H氧化成一分 子H2O,生成3分子ATP,而FADH2参与的递氢体系则生成2分子ATP,再加上三羧酸循环中有一次底物磷酸化产生一分子ATP,那么,一分子 CH2CO?SCoA参与三羧酸循环,直至循环终末共生成12分子ATP。 ③乙酰CoA中乙酰基的碳原子,乙酰CoA进入循环,与四碳受体分子草酰乙酸缩合,生成六碳的柠檬酸,在三羧酸循环中有二次脱羧生成2分 子CO2,与进入循环的二碳乙酰基的碳原子数相等,但是,以CO2方式失去的碳并非来自乙酰基的两个碳原子,而是来自草酰乙酸。 ④三羧酸循环的中间产物,从理论上讲,可以循环不消耗,但是由于循环中的某些组成成分还可参与合成其他物质,而其他物质也可不断通过 多种途径而生成中间产物,所以说三羧酸循环组成成分处于不断更新之中。 例如 草楚酰乙酸——→天门冬氨酸 α-酮戊二酸——→谷氨酸 草酰乙酸——→丙酮酸——→丙氨酸