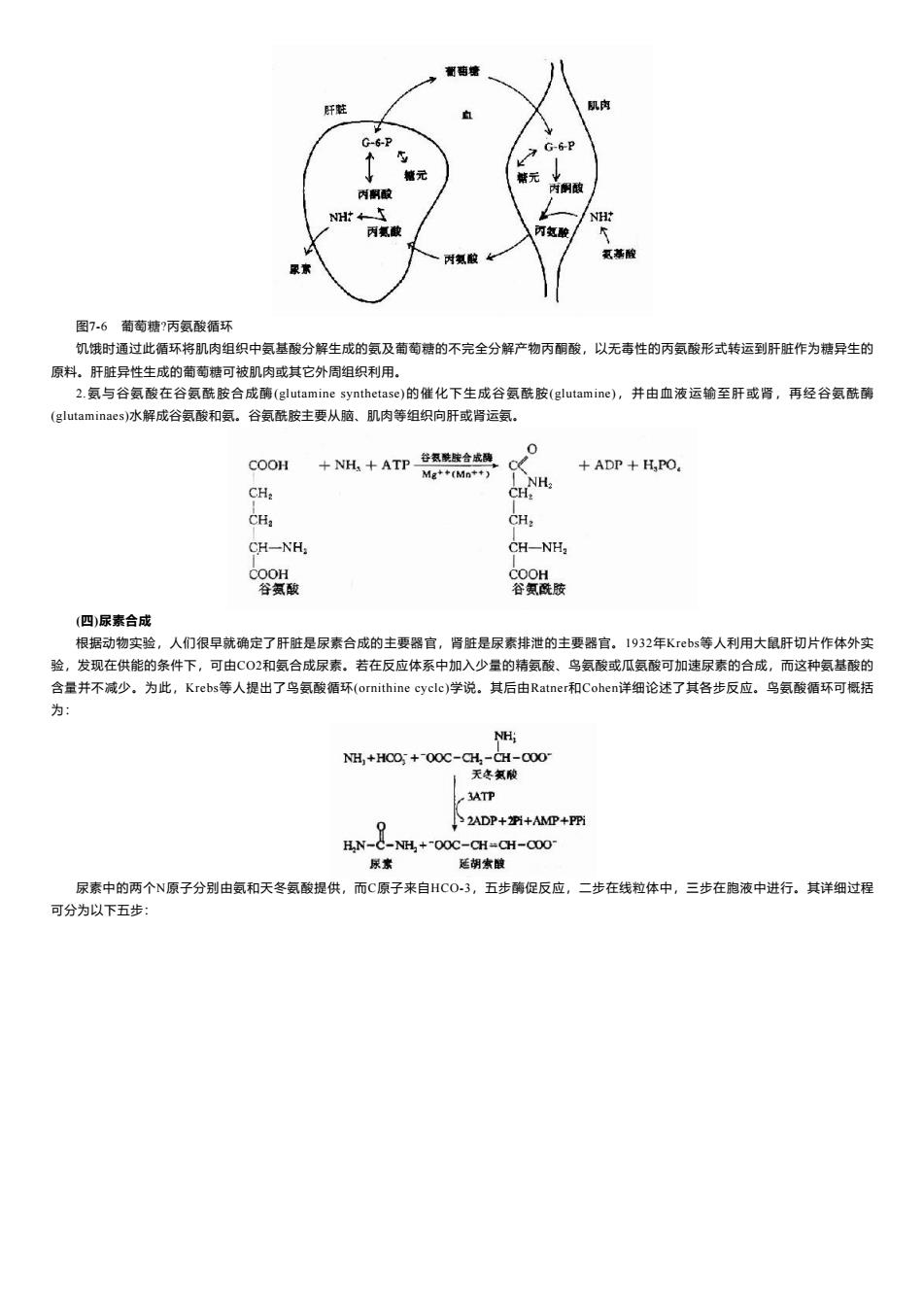
萄塘 肝鞋 肌肉 G-6-P G-6-P 个 辙元 储元 内酮酸 内酮酸 NH← NH 丙氨酸 丙氨酸 时氨酸女 氢善酸 图7-6葡萄糖?丙氨酸循环 饥饿时通过此循环将肌肉组织中氨基酸分解生成的氨及萄萄糖的不完全分解产物丙酮酸,以无毒性的丙氨酸形式转运到肝脏作为糖异生的 原料。肝脏异性生成的萄萄糖可被肌肉或其它外周组织利用。 2.氨与谷氨酸在谷氨酰胺合成酶(glutamine synthetase)的催化下生成谷氨酰胺(glutamine),并由血液运输至肝或肾,再经谷氨酰酶 (glutaminaes)水解成谷氨酸和氨。谷氨酰胺主要从脑、肌肉等组织向肝或肾运氨。 0 COOH 十NH十ATP谷氨酰合成 .ce +ADP H,PO, Mg**(Mn++) NH CH: CH: CH CH: CH-NH, CH-NH2 COOH COOH 谷氨酸 谷氨酰胺 (四)尿素合成 根据动物实验,人们很早就确定了肝脏是尿素合成的主要器官,肾脏是尿素排泄的主要器官。1932年Kbs等人利用大鼠肝切片作体外实 验,发现在供能的条件下,可由C02和氨合成尿素。若在反应体系中加入少量的精氨酸、鸟氨酸或瓜氨酸可加速尿素的合成,而这种氨基酸的 含量并不减少。为此,Krebs等人提出了鸟氨酸循环(ornithine cyclc)学说。其后由Ratner和Cohen详细论述了其各步反应。鸟氨酸循环可概括 为: NH; NH,+HCO +00C-CH,-CH-000- 天冬氨酸 3ATP 2ADP++AMP+P阳 H.N-C-NH +-00C-CH=CH-C0O- 尿家 延胡索皱 尿素中的两个N原子分别由氨和天冬氨酸提供,而C原子来自HCO-3,五步酶促反应,二步在线粒体中,三步在胞液中进行。其详细过程 可分为以下五步:
图7-6 葡萄糖?丙氨酸循环 饥饿时通过此循环将肌肉组织中氨基酸分解生成的氨及葡萄糖的不完全分解产物丙酮酸,以无毒性的丙氨酸形式转运到肝脏作为糖异生的 原料。肝脏异性生成的葡萄糖可被肌肉或其它外周组织利用。 2.氨与谷氨酸在谷氨酰胺合成酶(glutamine synthetase)的催化下生成谷氨酰胺(glutamine),并由血液运输至肝或肾,再经谷氨酰酶 (glutaminaes)水解成谷氨酸和氨。谷氨酰胺主要从脑、肌肉等组织向肝或肾运氨。 (四)尿素合成 根据动物实验,人们很早就确定了肝脏是尿素合成的主要器官,肾脏是尿素排泄的主要器官。1932年Krebs等人利用大鼠肝切片作体外实 验,发现在供能的条件下,可由CO2和氨合成尿素。若在反应体系中加入少量的精氨酸、鸟氨酸或瓜氨酸可加速尿素的合成,而这种氨基酸的 含量并不减少。为此,Krebs等人提出了鸟氨酸循环(ornithine cyclc)学说。其后由Ratner和Cohen详细论述了其各步反应。鸟氨酸循环可概括 为: 尿素中的两个N原子分别由氨和天冬氨酸提供,而C原子来自HCO-3,五步酶促反应,二步在线粒体中,三步在胞液中进行。其详细过程 可分为以下五步:

ADP 0 H0-C-0+0p-0 ADP 0 HO-C-OPO+:N 琉基碎酸 2 - O-C-NH, 氨基甲酸 0 P OP-0-C-NH, 氨基甲酰磷酸 图7-7CPS?I作用机理 1.氨基甲酰磷酸的合成 氨基甲酰磷酸(carbamylphosphate)是在Mg+、ATP及N?乙酰谷氨酸(N?acetyl glutamic acid,.AGA)存在的情况下,由氨基甲酰磷酸合成酶 I(carbamyl phosphate synthetasel,.CPS?I)催化NH3和HCO·3在肝细胞线粒体中合成。 NH,HCO+2ATP +H:O- 氛基甲酰磷酸 合成德【M一→ OH NH,-C-0 OH +H,PO,+2ADP 真核细胞中有两种CPS::(1)线粒体CPS?-I利用游离NH3为氨源合成氨基甲酰磷酸,参与尿素合成。(2)胞液CPS-Ⅱ,利用谷氨酰胺作N源, 参与嘧啶的从头合成。 CPS-I催化的反应包括下述三步(图7-7)。 (I)ATP活化HCO-3生成ADP和羰基硫酸(carbonyl phosphate) (2)NH2与羰基硫酸作用替代硫酸根,生成氨基甲酸(carbamate)和Pi。 (3)第2个ATP对氨甲酸磷酸化,生成氨基甲酰磷酸和ADP。 此反应是不可逆的,消耗2分子ATP。CPS?1是一种变构酶,AGA是此酶变构激活剂。由乙酰C0A和谷氨酸缩合而成 COOI CH-CO--SCoA+谷氢酸→CH,CO-NHCH iCoA SH CH; HOOC-CH AGA 肝细胞线粒体中谷氨酸脱氢酶和氨基甲酰磷酸合成酶I催化的反应是紧密偶联的。谷氨酸脱氢酶催化谷氨酸氧化脱氨,生成的产物有H3和 NADH+H+。NADH经NADH氧化呼吸链传递氧化生成H2O,释放出来的能量用于ADP磷酸化生成ATP。因此谷氨酸脱氢酶催化反应不仅为氨 基甲酰磷酸的合成提供了底物NH3,同时也提供了该反应所需要的能量ATP。氨基甲酰磷酸合成酶I将有毒的氨转变成氨基甲酰磷酸,反应中生 成的ADP又是谷氨酸脱氢酶的变构激活剂,促进谷氨酸进一步氧化脱氨。这种紧密偶联有利于迅速将氨固定在肝细胞线粒体内,防止氨逸出线 粒体进入细胞浆,进而透过细胞膜进入血液,引起血氨升高。 2.瓜氨酸(citrulline)的生成: 乌氨酸氨基甲酰转移酶(ornithinetranscarbamoylase)存在于线粒体中,通常与CPS·I形成酶的复合物催化氨基甲酰磷酸转甲酰基给鸟氨酸 生成瓜氨酸。(注意:鸟氨酸,瓜氨酸均非标准α-氨基酸,不出现在蛋白质中)。此反应在线粒体内进行,而鸟氨酸在胞液中生成,所以必需通 过一特异的穿棱系统进入线粒体内
图7-7 CPS?Ⅰ 作用机理 1.氨基甲酰磷酸的合成 氨基甲酰磷酸(carbamylphosphate)是在Mg++、ATP及N?乙酰谷氨酸(N?acetyl glutamic acid,AGA)存在的情况下,由氨基甲酰磷酸合成酶 I(carbamyl phosphate synthetaseI, CPS?I)催化NH3和HCO-3在肝细胞线粒体中合成。 真核细胞中有两种CPS:(1)线粒体CPS?-Ⅰ利用游离NH3为氮源合成氨基甲酰磷酸,参与尿素合成。(2)胞液CPS-Ⅱ,利用谷氨酰胺作N源, 参与嘧啶的从头合成。 CPS-Ⅰ催化的反应包括下述三步(图7-7)。 (1)ATP活化HCO-3生成ADP和羰基硫酸(carbonyl phosphate) (2)NH2与羰基硫酸作用替代硫酸根,生成氨基甲酸(carbamate)和Pi。 (3)第2个ATP对氨甲酸磷酸化,生成氨基甲酰磷酸和ADP。 此反应是不可逆的,消耗2分子ATP。CPS?1是一种变构酶,AGA是此酶变构激活剂。由乙酰CoA和谷氨酸缩合而成。 肝细胞线粒体中谷氨酸脱氢酶和氨基甲酰磷酸合成酶I催化的反应是紧密偶联的。谷氨酸脱氢酶催化谷氨酸氧化脱氨,生成的产物有NH3和 NADH+H+。NADH经NADH氧化呼吸链传递氧化生成H2O,释放出来的能量用于ADP磷酸化生成ATP。因此谷氨酸脱氢酶催化反应不仅为氨 基甲酰磷酸的合成提供了底物NH3,同时也提供了该反应所需要的能量ATP。氨基甲酰磷酸合成酶I将有毒的氨转变成氨基甲酰磷酸,反应中生 成的ADP又是谷氨酸脱氢酶的变构激活剂,促进谷氨酸进一步氧化脱氨。这种紧密偶联有利于迅速将氨固定在肝细胞线粒体内,防止氨逸出线 粒体进入细胞浆,进而透过细胞膜进入血液,引起血氨升高。 2.瓜氨酸(citrulline)的生成: 乌氨酸氨基甲酰转移酶(ornithinetranscarbamoylase)存在于线粒体中,通常与CPS-I形成酶的复合物催化氨基甲酰磷酸转甲酰基给鸟氨酸 生成瓜氨酸。(注意:鸟氨酸,瓜氨酸均非标准α-氨基酸,不出现在蛋白质中)。此反应在线粒体内进行,而鸟氨酸在胞液中生成,所以必需通 过一特异的穿棱系统进入线粒体内