
别去碱基的数目 0 CATCATCAT CAT CAT CAT CAT 1 CAT CTC ATCATC ATC ATC ATC A 2 CAT CTC ACA TCA TCATCA TCA A T CAT CTC ACA TAT CATCAT CAT A T 当删去一个碱基A时,从这一点以后的密码就发生了差错。删去两个碱基时,情形 也如此。但是别去三个碱基时,情况就不同了。最先也形成几组错误的密码子,但以后又 恢复正常。前面两类突变往往使基因产物全部失去活力,而第三种突变类型使基因产物仍 具有一定活力。这只能用遗传密码是三联体这个事实来加以解释。 2.遗传密码的主要特征: (1)密码的无标点性即两个密码子之间没有任何起标点符号作用的密码子加以隔 开。因此要正确阅读密码必须按一定的读码框架((reading frame),从一个正确的起点开始, 一个不漏地挨若读下去,直至碰到终止信号为止。若插入(insertion)或刑去(deletion)一个 碱基,就会使这以后的读码发生错误,这称移码(rame-shif)。由于移码引起的突变称移 码突变(frame-shift mutation)。 (2)一般情形下遗传密码是不重叠(non-overlapping)的是指每三个碱基编码一个 氨基酸,碱基不重复使用。即 ABC DEF GHI JKL aa.aa an aa. 目前已经证明,在绝大多数生物中读码规则是不重叠的。但是在少数大肠杆菌噬菌体 (如R、QB等)的RNA基因组中,部分基因的遗传密码却是重叠的。 (3)密码的简并性(degeneracy)是指大多数氨基酸都可以具有几组不同的密码子 (见表-2)。如UUA、UUG、CUU、CUC、CUA、CUG六组密码子都编码亮氨酸。编 码同一个氨基酸的一组密码称为同义密码子。只有色氨酸和甲硫氨酸仅有一个密码子。 密码的简并性具有重要的生物学意义。它可以减少有害的突变。一方面,如果每个氨 基酸只有一个密码子,20组密码子就可以应付20种氨基酸的编码了,那么剩下的4组密 码子都将会导致肽链合成的终止。由于突变而引起的肽链合成终止的频率也会大大提高。 这样合成出来的残缺不全的多肽往往不具有生物活力。 332
332 删去碱基的数目 0 CAT CAT CAT CAT CAT CAT CAT 1 CAT CTC ATC ATC ATC ATC ATC ↓ A 2 CAT CTC ACA TCA TCA TCA TCA ↓ ↓ A T 3 CAT CTC ACA TAT CAT CAT CAT ↓ ↓ ↓ A T C 当删去一个碱基 A 时,从这一点以后的密码就发生了差错。删去两个碱基时,情形 也如此。但是删去三个碱基时,情况就不同了。最先也形成几组错误的密码子,但以后又 恢复正常。前面两类突变往往使基因产物全部失去活力,而第三种突变类型使基因产物仍 具有一定活力。这只能用遗传密码是三联体这个事实来加以解释。 2.遗传密码的主要特征: (1)密码的无标点性 即两个密码子之间没有任何起标点符号作用的密码子加以隔 开。因此要正确阅读密码必须按一定的读码框架(reading frame),从一个正确的起点开始, 一个不漏地挨着读下去,直至碰到终止信号为止。若插入(insertion)或删去(deletion)一个 碱基,就会使这以后的读码发生错误,这称移码(frame-shift)。由于移码引起的突变称移 码突变(frame-shift mutation)。 (2)一般情形下遗传密码是不重叠(non-overlapping)的 是指每三个碱基编码一个 氨基酸,碱基不重复使用。即 ABC DEF GHI JKL ↓ ↓ ↓ ↓ aa1 aa2 aa3 aa4 目前已经证明,在绝大多数生物中读码规则是不重叠的。但是在少数大肠杆菌噬菌体 (如 R17、Qβ等)的 RNA 基因组中,部分基因的遗传密码却是重叠的。 (3)密码的简并性(degeneracy) 是指大多数氨基酸都可以具有几组不同的密码子 (见表 11-2)。如 UUA、UUG、CUU、CUC、CUA、CUG 六组密码子都编码亮氨酸。编 码同一个氨基酸的一组密码称为同义密码子。只有色氨酸和甲硫氨酸仅有一个密码子。 密码的简并性具有重要的生物学意义。它可以减少有害的突变。一方面,如果每个氨 基酸只有一个密码子,20 组密码子就可以应付 20 种氨基酸的编码了,那么剩下的 44 组密 码子都将会导致肽链合成的终止。由于突变而引起的肽链合成终止的频率也会大大提高。 这样合成出来的残缺不全的多肽往往不具有生物活力
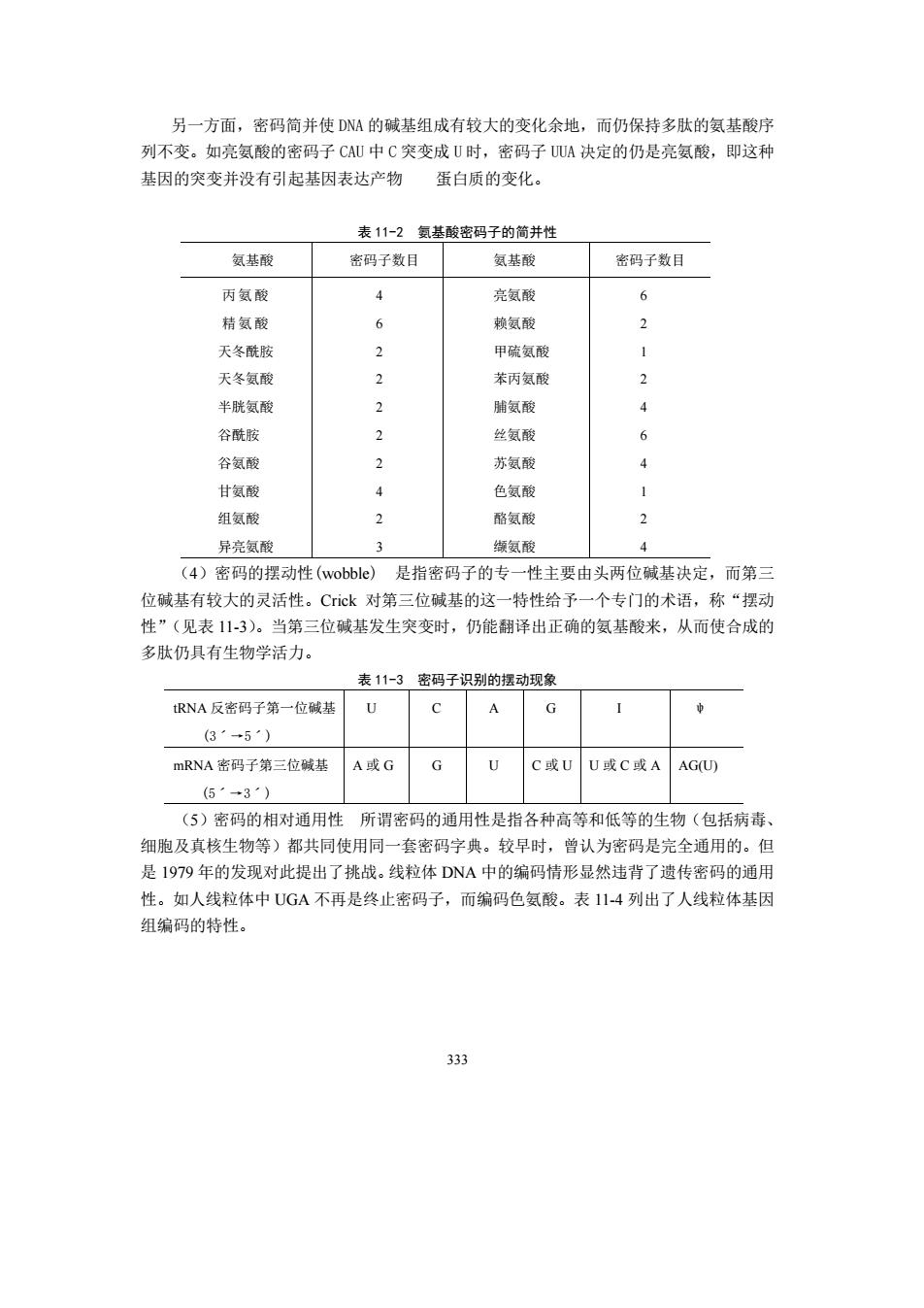
另一方面,密码简并使DNA的碱基组成有较大的变化余地,而仍保持多肽的氨基酸序 列不变。如亮氨酸的密码子CAU中C突变成U时,密码子UA决定的仍是亮氨酸,即这种 基因的突变并没有引起基因表达产物 蛋白质的变化。 表11-2氨基酸密码子的简并性 氨基酸 密码子数目 氨基酸 密码子数目 丙氨酸 4 亮氨酸 6 桔氨酸 6 赖氨酸 2 天冬酰脑 2 甲硫氨酸 天冬氨酸 苯丙氨酸 2 半胱氨酸 2 脯氨酸 4 谷酰胺 2 丝氨酸 6 谷氨酸 苏氨酸 甘氨酸 ¥ 色氨酸 1 组氨酸 2 酪氨酸 2 异亮氨酸 3 缬氨酸 4 (4)密码的摆动性(wobbIe) 是指密码子的专一性主要由头两位碱基决定,而第 位碱基有较大的灵活性。Cck对第三位碱基的这一特性给予一个专门的术语,称“摆动 性”(见表11-3)。当第三位碱基发生突变时,仍能翻译出正确的氨基酸来,从而使合成的 多肽仍具有生物学活力。 表11-3密码子识别的摆动现象 RNA反密码子第一位碱热 (3·→5·) mRNA密码子第三位碱基A或GG UC或UU或C或AAGU 5·3·) (5)密码的相对通用性所谓密码的通用性是指各种高等和低等的生物(包括病毒、 细胞及真核生物等)都共同使用同一套密码字典。较早时,曾认为密码是完全通用的。但 是1979年的发现对此提出了挑战。线粒体DNA中的编码情形显然违背了遗传密码的通用 性。如人线粒体中UG不再是终止密码子,而编码色氨酸。表114列出了人线粒体基因 组编码的特性。 333
333 另一方面,密码简并使 DNA的碱基组成有较大的变化余地,而仍保持多肽的氨基酸序 列不变。如亮氨酸的密码子 CAU中 C突变成 U时,密码子 UUA决定的仍是亮氨酸,即这种 基因的突变并没有引起基因表达产物 蛋白质的变化。 表 11-2 氨基酸密码子的简并性 氨基酸 密码子数目 氨基酸 密码子数目 丙氨酸 精氨酸 天冬酰胺 天冬氨酸 半胱氨酸 谷酰胺 谷氨酸 甘氨酸 组氨酸 异亮氨酸 4 6 2 2 2 2 2 4 2 3 亮氨酸 赖氨酸 甲硫氨酸 苯丙氨酸 脯氨酸 丝氨酸 苏氨酸 色氨酸 酪氨酸 缬氨酸 6 2 1 2 4 6 4 1 2 4 (4)密码的摆动性(wobble) 是指密码子的专一性主要由头两位碱基决定,而第三 位碱基有较大的灵活性。Crick 对第三位碱基的这一特性给予一个专门的术语,称“摆动 性”(见表 11-3)。当第三位碱基发生突变时,仍能翻译出正确的氨基酸来,从而使合成的 多肽仍具有生物学活力。 表 11-3 密码子识别的摆动现象 tRNA 反密码子第一位碱基 (3ˊ→5ˊ) U C A G I ψ mRNA 密码子第三位碱基 (5ˊ→3ˊ) A 或 G G U C 或 U U 或 C 或 A AG(U) (5)密码的相对通用性 所谓密码的通用性是指各种高等和低等的生物(包括病毒、 细胞及真核生物等)都共同使用同一套密码字典。较早时,曾认为密码是完全通用的。但 是 1979 年的发现对此提出了挑战。线粒体 DNA 中的编码情形显然违背了遗传密码的通用 性。如人线粒体中 UGA 不再是终止密码子,而编码色氨酸。表 11-4 列出了人线粒体基因 组编码的特性

表11-4人线粒体DW中密码编制特点 密码 “通用”密码 人线粒体密码 UGA 终止密码 Trp AGA Arg 终止密码 AGG g 终止密码 AUA lle 起始密码(Me或le) AUU Ile 起始密码(le) AUG 起始密码(Mct或fMct) 起始毫码(ct)】 酵母线粒体原生动物纤毛虫也有类似情形。 AGA, AGG不再是终止信号而编码精氨 酸。 所以,遗传密码具有相对的通用性。 (6)起始密码子和终止密码子在64种密码子中,AUG既是甲硫氨酸的密码子, 又是肽链合成的起始密码子。有三组密码子UAA,UAG,UGA不编码任何氨基酸而成为 肽链合成的终止密码子,又称无义密码子。 二、tRNA 在蛋白质合成中,tRNA是搬运活性氨基酸的工具。它将氨基酸按照mRNA链上的密 码子所决定的氨基酸顺序搬运到蛋白质合成的场所核糖体的特定部位。RNA是多肽 麟和mRNA之间的重要转换器(adaptor),每一种氨基酸可以有一种以上tRNA作为运载工 具,人们把携带相同氨基酸而反密码子不同的一组tRNA称为同功受体RNA((isoaccepting tRNAs))。RNA在识别密码子上的接头作用: RNA分子上与多肽合成有关的位点至少有四个: 1.3”端-CCA上的氨基酸接受位点 RNA分子的3·端的碱基顺序是-CCA,“活化的氨基酸的羧基连接到3·末端腺苷 的核糖3·OH上,形成氨酰RNA。 A 334
334 表 11-4 人线粒体 DNA中密码编制特点 密码 “通用” 密码 人线粒体密码 UGA AGA AGG AUA AUU AUG 终止密码 Arg Arg Ile Ile 起始密码(Met 或 fMet) Trp 终止密码 终止密码 起始密码(Met 或 Ile) 起始密码(Ile) 起始密码(Met) 酵母线粒体原生动物纤毛虫也有类似情形。AGA,AGG 不再是终止信号而编码精氨 酸。 所以,遗传密码具有相对的通用性。 (6)起始密码子和终止密码子 在 64 种密码子中,AUG 既是甲硫氨酸的密码子, 又是肽链合成的起始密码子。有三组密码子 UAA,UAG,UGA 不编码任何氨基酸而成为 肽链合成的终止密码子,又称无义密码子。 二、tRNA 在蛋白质合成中,tRNA 是搬运活性氨基酸的工具。它将氨基酸按照 mRNA 链上的密 码子所决定的氨基酸顺序搬运到蛋白质合成的场所 核糖体的特定部位。tRNA 是多肽 链和 mRNA 之间的重要转换器(adaptor),每一种氨基酸可以有一种以上 tRNA 作为运载工 具,人们把携带相同氨基酸而反密码子不同的一组 tRNA 称为同功受体 tRNA(isoaccepting tRNAs)。tRNA 在识别密码子上的接头作用: tRNA 分子上与多肽合成有关的位点至少有四个: 1. 3ˊ端-CCA 上的氨基酸接受位点 tRNA 分子的 3ˊ端的碱基顺序是-CCA,“活化¡±的氨基酸的羧基连接到 3ˊ末端腺苷 的核糖 3ˊ-OH 上,形成氨酰- tRNA

2.识别氨酰-RNA合成酶的位点 形成氨酰:RNA的反应是在氨酰RNA合成酶催化下完成的。这个反应需要三种底 物,即氨基酸、RNA和ATP。由ATP提供活化氨基酸所需要的能量。一种氨酰-RNA 合成班以识别一组同功受体RNA(最多达6个)。 3.核糖体识别位点 在核糖体内合成多肽链的过程中,多肽链通过tRNA暂时结合在核糖体的正确位置上, 直至合成终止后多肽链才从核糖体上脱下。RNA起着连接这条多肽链和核糖体的作用。 4.反密码子位点 在RNA链上有三个特定的碱基,组成一个反密码子,反码 子与密码子的方向相反。由这反密码子按碱基配对原则识别mRNA链上的密码子(见图 II3)。一种RNA分子常常能够识别一种以上的同义密码子,这是因为RNA分子上的反 密码子与密码子的配对具有摆动性,配对的摆动性是由RNA反密码子环的空间结构决定 的。反密码子5‘端的碱基处于L形tRNA的顶端,受到的碱基堆积力的束缚较小,因此 有较大的自由度。而且该位置的碱基常为修饰的碱基,如次黄票吟1,它可以和U、C、 A三种碱基配对,具有非凡的“阅读”能力。分析表明同义密码子的使用频率是不相同的, 它与细胞内RNA含量(即RNA的丰度)成正相关,含量高的同功受体RNA所对应的 密码子的使用频*总是最高 CC G 3 mRNA 图1-3密码子与反密码子之间的识别 三、rRNA与核糖体 核糖体是蛋白质合成的装配机,是由几十种蛋白质和几种RA组成的亚细胞颗粒,其 中蛋白质与RNA的重量比约为1:2,是tRNA、mRNA和蛋白质相互作用的场所。早在 1950年就有人将放射性同位素标记的氨基酸注射到小鼠体内,经短时间后,取出肝脏,制 335
335 2.识别氨酰-tRNA 合成酶的位点 形成氨酰-tRNA 的反应是在氨酰-tRNA 合成酶催化下完成的。这个反应需要三种底 物,即氨基酸、tRNA 和 ATP 。由 ATP 提供活化氨基酸所需要的能量。一种氨酰-tRNA 合成酶可以识别一组同功受体 tRNA(最多达 6 个)。 3.核糖体识别位点 在核糖体内合成多肽链的过程中,多肽链通过 tRNA 暂时结合在核糖体的正确位置上, 直至合成终止后多肽链才从核糖体上脱下。tRNA 起着连接这条多肽链和核糖体的作用。 4.反密码子位点 在 tRNA 链上有三个特定的碱基,组成一个反密码子,反密码 子与密码子的方向相反。由这反密码子按碱基配对原则识别 mRNA 链上的密码子(见图 11-3)。一种 tRNA 分子常常能够识别一种以上的同义密码子,这是因为 tRNA 分子上的反 密码子与密码子的配对具有摆动性,配对的摆动性是由 tRNA 反密码子环的空间结构决定 的。反密码子 5ˊ端的碱基处于 L 形 tRNA 的顶端,受到的碱基堆积力的束缚较小,因此 有较大的自由度。而且该位置的碱基常为修饰的碱基,如次黄嘌呤 I,它可以和 U、C 、 A 三种碱基配对,具有非凡的“阅读”能力。分析表明同义密码子的使用频率是不相同的, 它与细胞内 tRNA 含量(即 tRNA 的丰度)成正相关,含量高的同功受体 tRNA 所对应的 密码子的使用频率总是最高。 图 11-3 密码子与反密码子之间的识别 三、rRNA 与核糖体 核糖体是蛋白质合成的装配机,是由几十种蛋白质和几种 RNA组成的亚细胞颗粒,其 中蛋白质与 RNA 的重量比约为 1∶2,是 tRNA、mRNA 和蛋白质相互作用的场所。早在 1950 年就有人将放射性同位素标记的氨基酸注射到小鼠体内,经短时间后,取出肝脏,制

成匀浆,离心,分成核、线粒拉体、微粒体及上清等组分。发现微粒体中的放射性强度最高 再用去污剂(如脱氧胆酸)处理微粒体,将核糖体从内质网中分离出来,发现核糖体的放 射强度比微粒体的要高7倍。这就说明核糖体是合成蛋白质的部位。 核糖体是一个巨大的核糖体蛋白体。在原核细胞中,它可以游离形式存在,也可以与 mRNA结合形成串状的多核糖体。平均每个细胞约有2000个核糖体。真核细胞中的核糖 体既可游离存在,也可以与细胞内质树相结合,形成粗植内质网。每个真核细胞所含核糖 体的数目要多得多,为10-10个。线粒体、叶绿体及细胞核内也有自己的核糖体。表11-5 总结了不同生物核糖体的一些特性。 表11-5核糖体的某些特性 核糖体种类 亚基 RNA(分子量) 蛋白质分子数目 原核细胞核糖休 30s 16S(5.510 21 (以大肠杆茵为例 70s r5S(0.4103) 34 50s 235(11010) 40S 18S(7010) n30 真核细胞核糖体 -5s(0410) ∽50 60 128-295(140-180105) 核糖体含3种RNA和55种蛋白质,这些成分如何组装成具有活性的核糖体,是 个值得深入研究的问题。在1968年第一次完成了大肠杆菌核糖体小亚基由其RNA和蛋 白质在体外的重新组装。这个重组装的颗粒具有与30S亚基功能完全相同的蛋白质合成活 性。重组装只需16SNA和21种蛋白质,而不需要加入其它组分(如酶或特殊因子),表 明这是一个“自我组装”(self-assembly)的过程。所谓自我组装,是指进行组装所需要的 全部信息都在亚基结构里,其蛋白质和RNA都带有规定组装过程的信息。自我组装的驱 动力包括水性相互作用、氢键和离子相互作用,以及碱基堆叠之间的相互作用等。 讲一一步的研究发现,这个组装过程有一定顺序,即某种蛋白质的加入要先于其它蛋白 质的加入。而且各组分的加入是有协同作用的,即一种组分的加入加强了下一种组分的加 入。图11-4概略地表示出大肠杆菌核糖体的30S和50S亚基的组装过程。在30S亚基的 组装过程中,以16SRNA为骨架,先与15种蛋白质结合(实际上,这些蛋白质的结合也 有先后),形成21S颗粒。然后再加上其余6种蛋白质,最后组装成30S亚基。这个组装 过程,可能在16SRNA转录开始之后即行发生,因为在RNA链上结合最强的部位,都 集中在最先转录的5端。 大肠杆菌核糖体50S亚基的组装比较复杂,它包括两种(23S和5S)rRNA,而且蛋白 质数目也较多。先由23SRNA,5SRNA与约20种L蛋白结合,生成33S颗粒。然后再 加上其余蛋白质,组装成41S颗粒。最后在Mg2+(0C)和50℃下转变为50S亚基。 336
336 成匀浆,离心,分成核、线粒体、微粒体及上清等组分。发现微粒体中的放射性强度最高。 再用去污剂(如脱氧胆酸)处理微粒体,将核糖体从内质网中分离出来,发现核糖体的放 射强度比微粒体的要高 7 倍。这就说明核糖体是合成蛋白质的部位。 核糖体是一个巨大的核糖体蛋白体。在原核细胞中,它可以游离形式存在,也可以与 mRNA 结合形成串状的多核糖体。平均每个细胞约有 2 000 个核糖体。真核细胞中的核糖 体既可游离存在,也可以与细胞内质网相结合,形成粗糙内质网。每个真核细胞所含核糖 体的数目要多得多,为 10 6 ~10 7个。线粒体、叶绿体及细胞核内也有自己的核糖体。表 11-5 总结了不同生物核糖体的一些特性。 表 11-5 核糖体的某些特性 核糖体种类 亚基 rRNA(分子量) 蛋白质分子数目 原核细胞核糖体 (以大肠杆菌为例) 30S 70S 50S 16S(5.5 10 5) 5S(0.4 10 5) 23S(110 10 5) 21 34 真核细胞核糖体 40S 80S 60S 18S(∽70 10 5) 5S(0.4 10 5) 28-29S(140-180 10 5) ∽30 ∽50 核糖体含 3 种 rRNA 和 55 种蛋白质,这些成分如何组装成具有活性的核糖体,是一 个值得深入研究的问题。在 1968 年第一次完成了大肠杆菌核糖体小亚基由其 rRNA 和蛋 白质在体外的重新组装。这个重组装的颗粒具有与 30S 亚基功能完全相同的蛋白质合成活 性。重组装只需 16SrRNA 和 21 种蛋白质,而不需要加入其它组分(如酶或特殊因子),表 明这是一个“自我组装”(self-assembly)的过程。所谓自我组装,是指进行组装所需要的 全部信息都在亚基结构里,其蛋白质和 rRNA 都带有规定组装过程的信息。自我组装的驱 动力包括水性相互作用、氢键和离子相互作用,以及碱基堆叠之间的相互作用等。 进一步的研究发现,这个组装过程有一定顺序,即某种蛋白质的加入要先于其它蛋白 质的加入。而且各组分的加入是有协同作用的,即一种组分的加入加强了下一种组分的加 入。图 11-4 概略地表示出大肠杆菌核糖体的 30S 和 50S 亚基的组装过程。在 30S 亚基的 组装过程中,以 16SrRNA 为骨架,先与 15 种蛋白质结合(实际上,这些蛋白质的结合也 有先后),形成 21S 颗粒。然后再加上其余 6 种蛋白质,最后组装成 30S 亚基。这个组装 过程,可能在 16SrRNA 转录开始之后即行发生,因为在 rRNA 链上结合最强的部位,都 集中在最先转录的 5ˊ端。 大肠杆菌核糖体 50S 亚基的组装比较复杂,它包括两种(23S 和 5S)rRNA,而且蛋白 质数目也较多。先由 23SrRNA,5SrRNA 与约 20 种 L 蛋白结合,生成 33S 颗粒。然后再 加上其余蛋白质,组装成 41S 颗粒。最后在 Mg 2+(0℃)和 50℃下转变为 50S 亚基