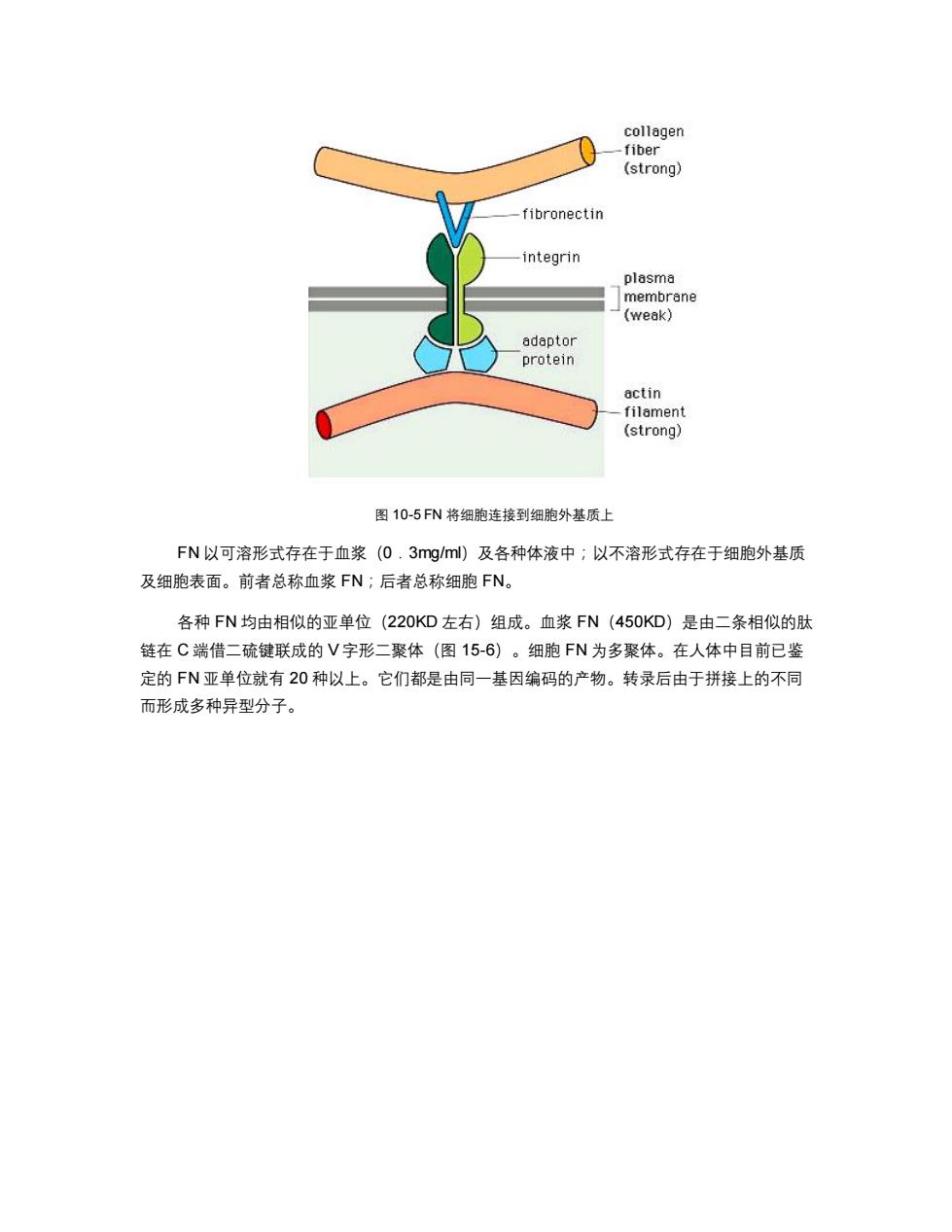
collagen fiber (strong) fibronectin integrin plasma membrane (weak) adaptor protein actin filament (strong) 图10-5FN将细胞连接到细胞外基质上 FN以可溶形式存在于血浆(0.3 mg/ml)及各种体液中;以不溶形式存在于细胞外基质 及细胞表面。前者总称血浆FN;后者总称细胞FN。 各种FN均由相似的亚单位(220KD左右)组成。血浆FN(450KD)是由二条相似的肽 链在C端借二硫键联成的V字形二聚体(图15-6)。细胞FN为多聚体。在人体中目前已鉴 定的FN亚单位就有20种以上。它们都是由同一基因编码的产物。转录后由于拼接上的不同 而形成多种异型分子
图 10-5 FN 将细胞连接到细胞外基质上 FN 以可溶形式存在于血浆(0.3mg/ml)及各种体液中;以不溶形式存在于细胞外基质 及细胞表面。前者总称血浆 FN;后者总称细胞 FN。 各种 FN 均由相似的亚单位(220KD 左右)组成。血浆 FN(450KD)是由二条相似的肽 链在 C 端借二硫键联成的 V 字形二聚体(图 15-6)。细胞 FN 为多聚体。在人体中目前已鉴 定的 FN 亚单位就有 20 种以上。它们都是由同一基因编码的产物。转录后由于拼接上的不同 而形成多种异型分子
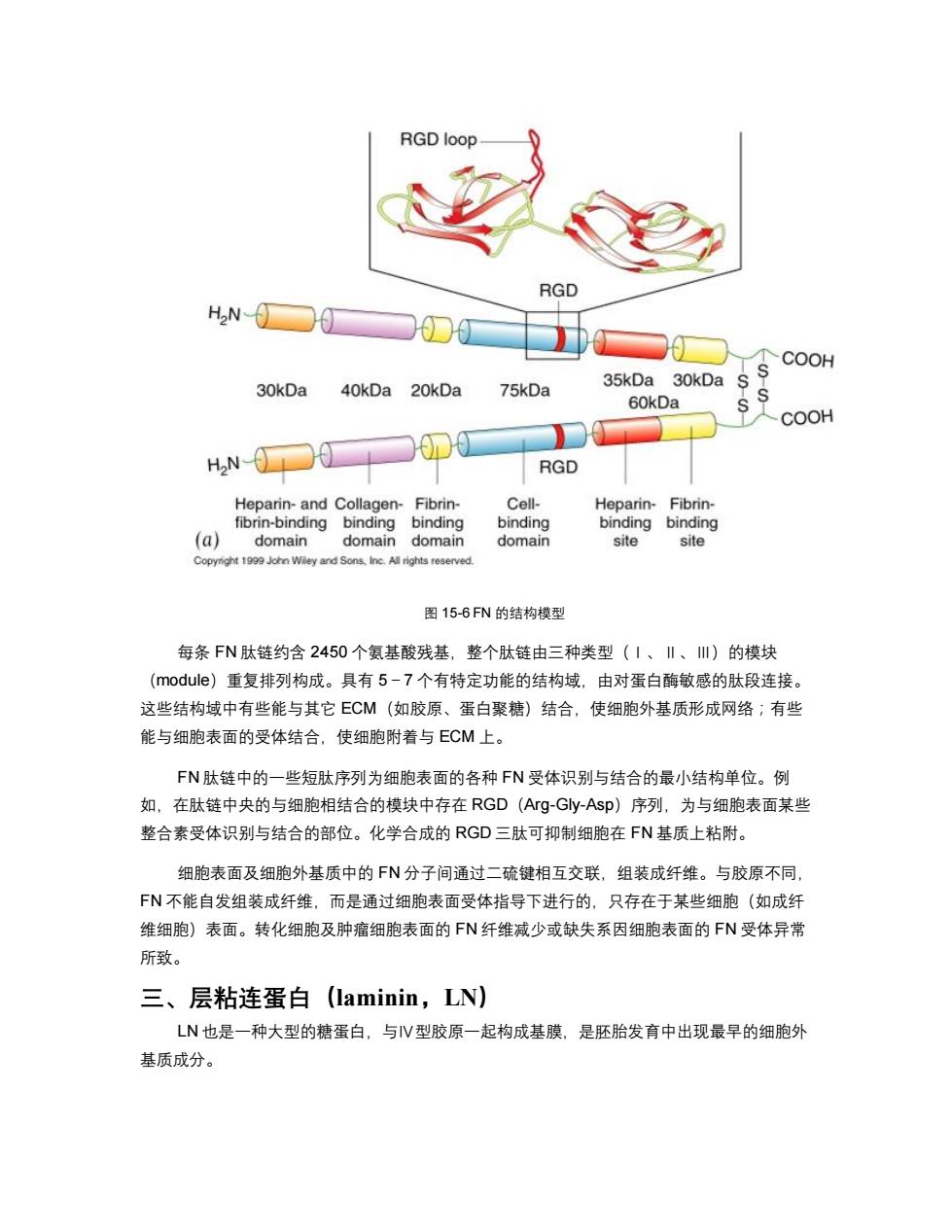
RGD loop RGD HN- TCOOH 30kDa 75kDa 35kDa 30kDa 40kDa 20kDa 60kDa COOH H2N RGD Heparin-and Collagen-Fibrin- Cell- Heparin-Fibrin- fibrin-binding binding binding binding binding binding (a) domain domain domain domain site site Copynight 1999 John Wley and Sons,Inc.All rights reserved. 图15-6FN的结构模型 每条FN肽链约含2450个氨基酸残基,整个肽链由三种类型(1、‖、Ⅲ)的模块 (module)重复排列构成。具有5-7个有特定功能的结构域,由对蛋白酶敏感的肽段连接。 这些结构域中有些能与其它ECM(如胶原、蛋白聚糖)结合,使细胞外基质形成网络;有些 能与细胞表面的受体结合,使细胞附着与ECM上。 FN肽链中的一些短肽序列为细胞表面的各种FN受体识别与结合的最小结构单位。例 如,在肽链中央的与细胞相结合的模块中存在RGD(Arg-Gly-Asp)序列,为与细胞表面某些 整合素受体识别与结合的部位。化学合成的RGD三肽可抑制细胞在FN基质上粘附。 细胞表面及细胞外基质中的FN分子间通过二硫键相互交联,组装成纤维。与胶原不同 FN不能自发组装成纤维,而是通过细胞表面受体指导下进行的,只存在于某些细胞(如成纤 维细胞)表面。转化细胞及肿瘤细胞表面的FN纤维减少或缺失系因细胞表面的FN受体异常 所致。 三、层粘连蛋白(laminin,LN) LN也是一种大型的糖蛋白,与V型胶原一起构成基膜,是胚胎发育中出现最早的细胞外 基质成分
图 15-6 FN 的结构模型 每条 FN 肽链约含 2450 个氨基酸残基,整个肽链由三种类型(Ⅰ、Ⅱ、Ⅲ)的模块 (module)重复排列构成。具有 5-7 个有特定功能的结构域,由对蛋白酶敏感的肽段连接。 这些结构域中有些能与其它 ECM(如胶原、蛋白聚糖)结合,使细胞外基质形成网络;有些 能与细胞表面的受体结合,使细胞附着与 ECM 上。 FN 肽链中的一些短肽序列为细胞表面的各种 FN 受体识别与结合的最小结构单位。例 如,在肽链中央的与细胞相结合的模块中存在 RGD(Arg-Gly-Asp)序列,为与细胞表面某些 整合素受体识别与结合的部位。化学合成的 RGD 三肽可抑制细胞在 FN 基质上粘附。 细胞表面及细胞外基质中的 FN 分子间通过二硫键相互交联,组装成纤维。与胶原不同, FN 不能自发组装成纤维,而是通过细胞表面受体指导下进行的,只存在于某些细胞(如成纤 维细胞)表面。转化细胞及肿瘤细胞表面的 FN 纤维减少或缺失系因细胞表面的 FN 受体异常 所致。 三、层粘连蛋白(laminin,LN) LN 也是一种大型的糖蛋白,与Ⅳ型胶原一起构成基膜,是胚胎发育中出现最早的细胞外 基质成分