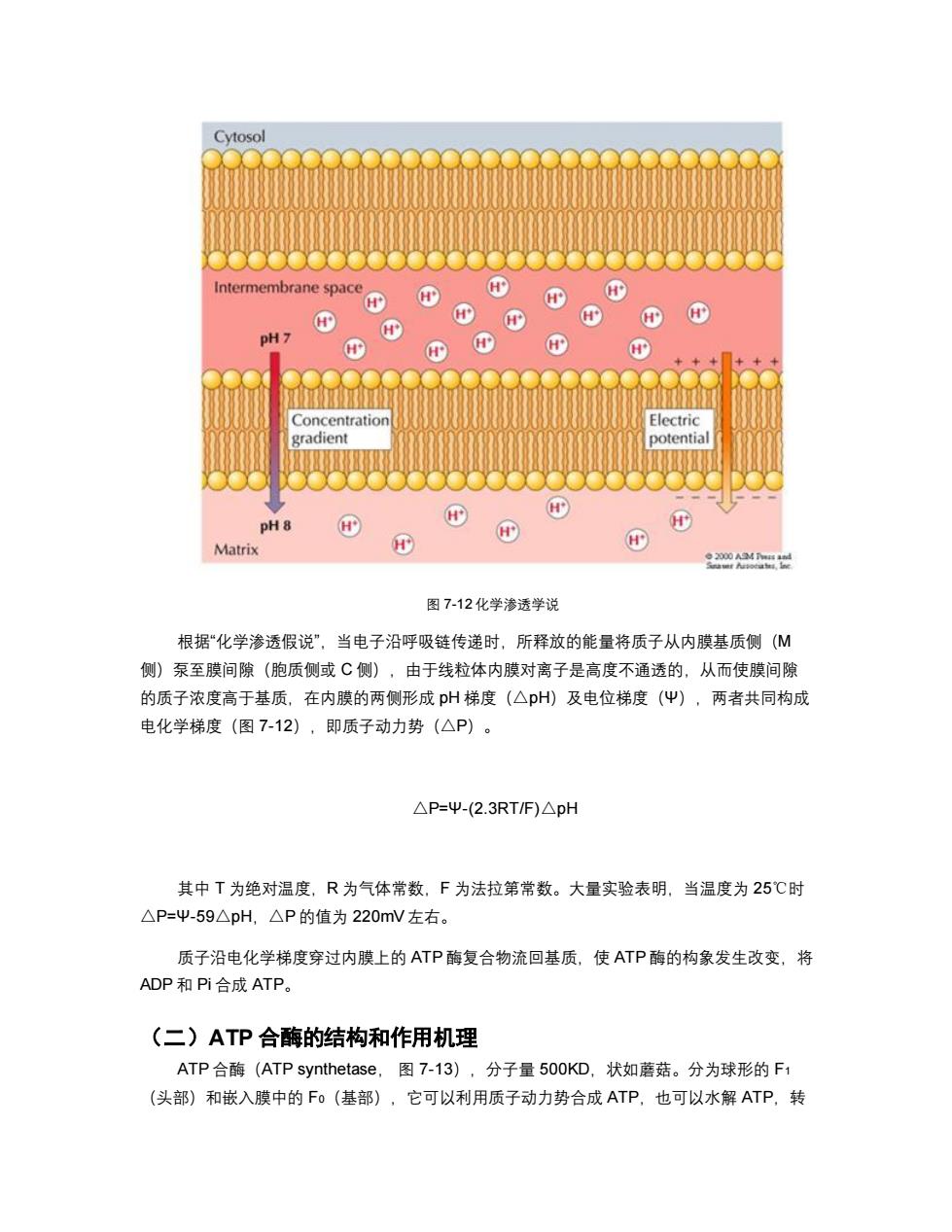
Cytosol Intermembrane space H H' H' PH7 H' Concentration Electric gradient potential pH8 H H' Matrix 22 图7-12化学渗透学说 根据“化学渗透假说”,当电子沿呼吸链传递时,所释放的能量将质子从内膜基质侧(M 侧)泵至膜间隙(胞质侧或C侧),由于线粒体内膜对离子是高度不通透的,从而使膜间隙 的质子浓度高于基质,在内膜的两侧形成p州梯度(△pH)及电位梯度(Ψ),两者共同构成 电化学梯度(图7-12),即质子动力势(△P)。 △P=Ψ-(2.3RTF)△pH 其中T为绝对温度,R为气体常数,F为法拉第常数。大量实验表明,当温度为25℃时 △P=Ψ-59△pH,△P的值为220mV左右。 质子沿电化学梯度穿过内膜上的ATP酶复合物流回基质,使ATP酶的构象发生改变,将 ADP和Pi合成ATP。 (二)ATP合酶的结构和作用机理 ATP合酶(ATP synthetase,图7-13),分子量500KD,状如蘑菇。分为球形的F1 (头部)和嵌入膜中的F0(基部),它可以利用质子动力势合成ATP,也可以水解ATP,转
图 7-12 化学渗透学说 根据“化学渗透假说”,当电子沿呼吸链传递时,所释放的能量将质子从内膜基质侧(M 侧)泵至膜间隙(胞质侧或 C 侧),由于线粒体内膜对离子是高度不通透的,从而使膜间隙 的质子浓度高于基质,在内膜的两侧形成 pH 梯度(△pH)及电位梯度(Ψ),两者共同构成 电化学梯度(图 7-12),即质子动力势(△P)。 △P=Ψ-(2.3RT/F)△pH 其中 T 为绝对温度,R 为气体常数,F 为法拉第常数。大量实验表明,当温度为 25℃时 △P=Ψ-59△pH,△P 的值为 220mV 左右。 质子沿电化学梯度穿过内膜上的 ATP 酶复合物流回基质,使 ATP 酶的构象发生改变,将 ADP 和 Pi 合成 ATP。 (二)ATP 合酶的结构和作用机理 ATP 合酶(ATP synthetase, 图 7-13),分子量 500KD,状如蘑菇。分为球形的 F1 (头部)和嵌入膜中的 F0(基部),它可以利用质子动力势合成 ATP,也可以水解 ATP,转

运质子,属于F型质子泵。每个肝细胞线粒体通常含15000个ATP合酶、每个酶每秒钟可产 生100个ATP。 F1由5种多肽组成a3β3Yδε复合体,具有三个ATP合成的催化位点(每个B亚基具有 个)。a和B单位交替排列,状如桔瓣。Y贯穿α邱复合体(相当于发电机的转子),并与Fo 接触,ε帮助Y与Fo结合。δ与Fo的两个b亚基形成固定a邱复合体的结构(相当于发电机的 定子)。 Rotation H 图7-13ATP合酶的结构(引自Lodish等1999) F0由三种多肽组成b2c12复合体,嵌入内膜,12个c亚基组成一个环形结构,具有质子 通道,可使质子由膜间隙流回基质。 1979年代Boyer P提出构象耦联假说,一些有力的实验证据使这一学说得到广泛的认 可。其要点如下: 1·ATP酶利用质子动力势,产生构象的改变,改变与底物的亲和力,催化ADP与Pi形 成ATP(图7-14)。 2,F1具有三个催化位点,但在特定的时间,三个催化位点的构象不同、因而与核苷酸的 亲和力不同。在L构象(loose),ADP、Pi与酶疏松结合在一起;在T构象((tight)底物 (ADP、Pi)与酶紧密结合在一起,在这种情况下可将两者加合在一起;在O构象(open) ATP与酶的亲和力很低,被释放出去
运质子,属于 F 型质子泵。每个肝细胞线粒体通常含 15000 个 ATP 合酶、每个酶每秒钟可产 生 100 个 ATP。 F1由 5 种多肽组成 α3β3γδε 复合体,具有三个 ATP 合成的催化位点(每个 β 亚基具有一 个)。α 和 β 单位交替排列,状如桔瓣。γ 贯穿 αβ 复合体(相当于发电机的转子),并与 F0 接触,ε 帮助 γ 与 F0结合。δ 与 F0的两个 b 亚基形成固定 αβ 复合体的结构(相当于发电机的 定子)。 图 7-13 ATP 合酶的结构(引自 Lodish 等 1999) F0由三种多肽组成 ab2c12复合体,嵌入内膜,12 个 c 亚基组成一个环形结构,具有质子 通道,可使质子由膜间隙流回基质。 1979 年代 Boyer P 提出构象耦联假说,一些有力的实验证据使这一学说得到广泛的认 可。其要点如下: 1.ATP 酶利用质子动力势,产生构象的改变,改变与底物的亲和力,催化 ADP 与 Pi 形 成 ATP(图 7-14)。 2.F1具有三个催化位点,但在特定的时间,三个催化位点的构象不同、因而与核苷酸的 亲和力不同。在 L 构象(loose),ADP、 Pi 与酶疏松结合在一起;在 T 构象(tight)底物 (ADP、 Pi)与酶紧密结合在一起,在这种情况下可将两者加合在一起;在 O 构象(open) ATP 与酶的亲和力很低,被释放出去
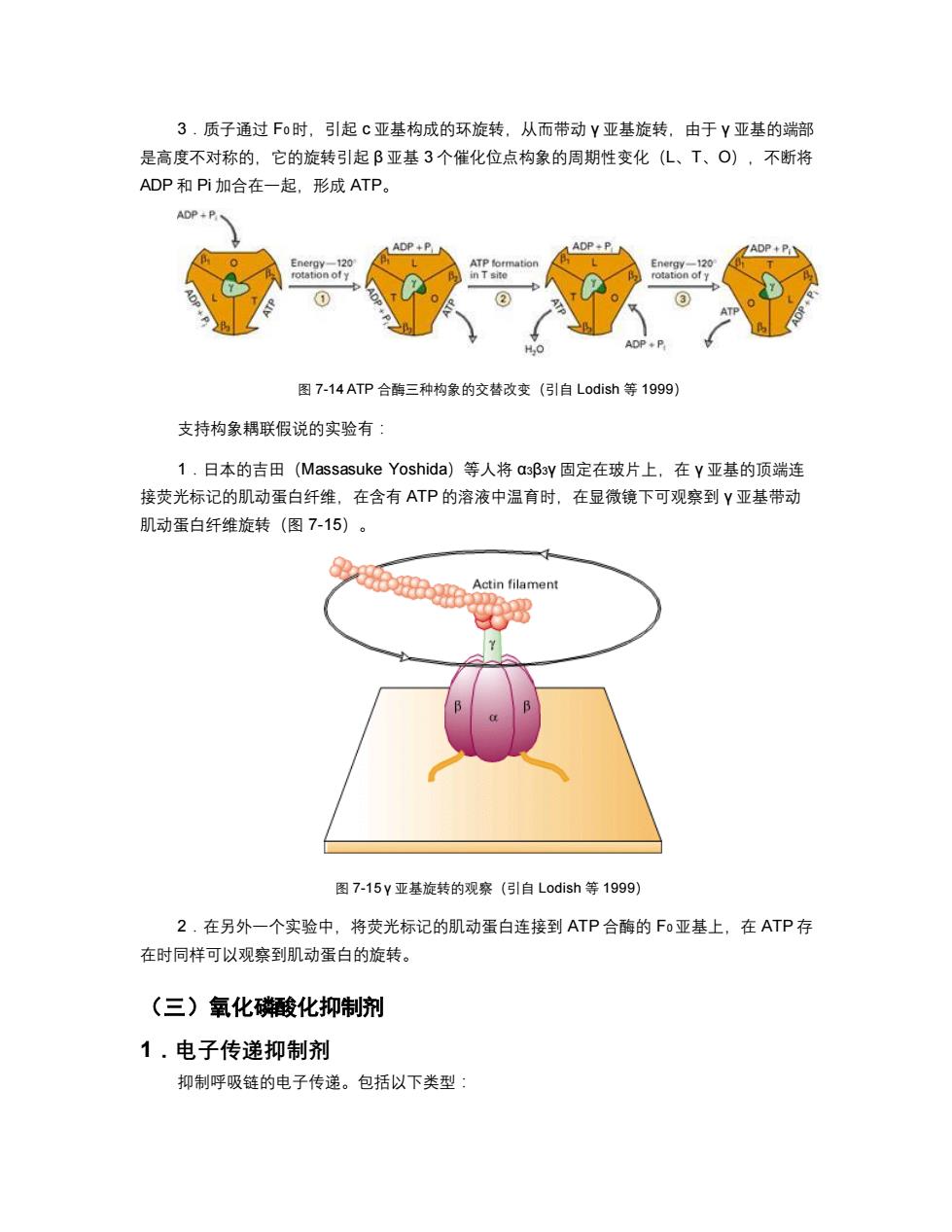
3.质子通过Fo时,引起c亚基构成的环旋转,从而带动Y亚基旋转,由于Y亚基的端部 是高度不对称的,它的旋转引起β亚基3个催化位点构象的周期性变化(L、T、O),不断将 ADP和Pi加合在一起,形成ATP。 ADP+P ADP+P ADP+PN ADP +P Energy-120 ATP formation Energy-120 rotation ofy in T site rotation of y H,O ADP◆P 图7-14ATP合酶三种构象的交替改变(引自Lodish等1999) 支持构象耦联假说的实验有: 1.日本的吉田(Massasuke Yoshida)等人将a3β3y固定在玻片上,在Y亚基的顶端连 接荧光标记的肌动蛋白纤维,在含有ATP的溶液中温育时,在显微镜下可观察到Y亚基带动 肌动蛋白纤维旋转(图7-15)。 Actin filament 图7-15y亚基旋转的观察(引自Lodish等1999) 2.在另外一个实验中,将荧光标记的肌动蛋白连接到ATP合酶的Fo亚基上,在ATP存 在时同样可以观察到肌动蛋白的旋转。 (三)氧化酸化抑制剂 1.电子传递抑制剂 抑制呼吸链的电子传递。包括以下类型:
3.质子通过 F0时,引起 c 亚基构成的环旋转,从而带动 γ 亚基旋转,由于 γ 亚基的端部 是高度不对称的,它的旋转引起 β 亚基 3 个催化位点构象的周期性变化(L、T、O),不断将 ADP 和 Pi 加合在一起,形成 ATP。 图 7-14 ATP 合酶三种构象的交替改变(引自 Lodish 等 1999) 支持构象耦联假说的实验有: 1.日本的吉田(Massasuke Yoshida)等人将 α3β3γ 固定在玻片上,在 γ 亚基的顶端连 接荧光标记的肌动蛋白纤维,在含有 ATP 的溶液中温育时,在显微镜下可观察到 γ 亚基带动 肌动蛋白纤维旋转(图 7-15)。 图 7-15 γ 亚基旋转的观察(引自 Lodish 等 1999) 2.在另外一个实验中,将荧光标记的肌动蛋白连接到 ATP 合酶的 F0亚基上,在 ATP 存 在时同样可以观察到肌动蛋白的旋转。 (三)氧化磷酸化抑制剂 1.电子传递抑制剂 抑制呼吸链的电子传递。包括以下类型:

①抑制NADH→CoQ的电子传递。如:阿米妥(amytal))、鱼藤酮(rotenone)、杀粉蝶素A (piericidin)。 ②抑制Cyt b-Cyt c1的电子传递。如:抗霉素A(antinomycin A)。 ③抑制细胞色素氧化酶→O2。如:CO、CN、NaN3、H2S。 电子传递抑制剂可用来研究呼吸链各组分的排列顺序,当呼吸链某一特定部位被抑制 后,底物一侧均为还原状态,氧一侧均为氧化态,可用分光光度计检测,因为电子传递链组分 氧化态和还原态具有不同的吸收峰。 2.磷酸化抑制剂 与Fo结合结合,阻断H通道,从而抑制ATP合成。如:寡霉素(oligomycin)、二环 己基碳化二亚胺(dicyclohexyl carbodiimide,DCC): 3.解偶联剂(uncoupler) 使氧化和磷酸化脱偶联,氧化仍可以进行,而磷酸化不能进行,解偶联剂为离子载体或 通道,能增大线粒体内膜对H的通透性,消除H梯度,因而无ATP生成,使氧化释放出来的 能量全部以热的形式散发。动物棕色脂肪组织和肌肉线粒体中有独特的解偶联蛋白 (uncoupling proteins,UCPs),与维持体温有关。常用解偶联剂主要有 质子载体:2,4-二硝基酚(DNP,图7-16),羰基-氰-对-三氟甲氧基苯肼(FCCP)。 质子通道:增温素(thermogenin)。 其它离子载体:如缬氨霉素。 某些药物:如过量的阿斯匹林也使氧化磷酸化部分解偶联,从而使体温升高。 NO2 NO 02N H" OH 图7-16DNP分子结构 四、线粒体的半自主性 1963年M.和S.Nass发现线粒体DNA(mtDNA)后,人们又在线粒体中发现了 RNA、DNA聚合酶、RNA聚合酶、tRNA、核糖体、氨基酸活化酶等进行DNA复制、转录和 蛋白质翻译的全套装备,说明线粒体具有独立的遗传体系
① 抑制 NADH→CoQ 的电子传递。如:阿米妥(amytal)、鱼藤酮(rotenone)、杀粉蝶素 A (piericidin)。 ② 抑制 Cyt b→Cyt c1的电子传递。如:抗霉素 A(antinomycin A)。 ③ 抑制细胞色素氧化酶→O2。如:CO、CN、NaN3、H2S。 电子传递抑制剂可用来研究呼吸链各组分的排列顺序,当呼吸链某一特定部位被抑制 后,底物一侧均为还原状态,氧一侧均为氧化态,可用分光光度计检测,因为电子传递链组分 氧化态和还原态具有不同的吸收峰。 2.磷酸化抑制剂 与 F0结合结合,阻断 H+通道,从而抑制 ATP 合成。如:寡霉素(oligomycin)、二环 己基碳化二亚胺(dicyclohexyl carbodiimide,DCC): 3.解偶联剂(uncoupler) 使氧化和磷酸化脱偶联,氧化仍可以进行,而磷酸化不能进行,解偶联剂为离子载体或 通道,能增大线粒体内膜对 H+的通透性,消除 H+梯度,因而无 ATP 生成,使氧化释放出来的 能量全部以热的形式散发。动物棕色脂肪组织和肌肉线粒体中有独特的解偶联蛋白 (uncoupling proteins, UCPs),与维持体温有关。常用解偶联剂主要有: 质子载体: 2,4-二硝基酚(DNP,图 7-16),羰基-氰-对-三氟甲氧基苯肼(FCCP)。 质子通道:增温素(thermogenin)。 其它离子载体:如缬氨霉素。 某些药物:如过量的阿斯匹林也使氧化磷酸化部分解偶联,从而使体温升高。 图 7-16 DNP 分子结构 四、线粒体的半自主性 1963 年 M. 和 S. Nass 发现线粒体 DNA(mtDNA)后,人们又在线粒体中发现了 RNA、DNA 聚合酶、RNA 聚合酶、tRNA、核糖体、氨基酸活化酶等进行 DNA 复制、转录和 蛋白质翻译的全套装备,说明线粒体具有独立的遗传体系