
球菌等 数种类如柄细菌分裂后产生一个有柄不运动利 napping division) 特殊的繁殖方式 细菌除无性繁殖外,电镜观察和遗传学研究己证明细菌存在着有性接 合。然而细菌有性接合较少,以无性繁殖为 图1.22细菌细胞的裂殖, 示正在分裂, ↑↑示己经分裂 (引自谢念铭主编《医学细菌电镜图谱》,人民卫生出版社,1994) (二)、细菌的群体特征 1、细菌菌落特征 细菌在周体培养基上生长发有,几天内即可由一个或几个细茵分裂繁 聚集在 起形成肉眼可见的群 菌落是由 个细菌菌体生长 在微生物的纯种分离中也可以挑志 、 图1·23细菌菌结 霸6 正面观: 扁平2.隆起3.低凸起4.高凸起5.脐状6.草 相状7到斗状 表面结构、形态及边缘:8.圆形、边缘完整9。不规则、边缘波 浪10.不规则、颗粒状、边缘叶状11.规划、放射状、边缘星 叶状12.规则、边缘呈扇边状13.规则、边缘呈齿状14.规则 跨@ 有同心环、边缘完整15.不规则、似毛毯状16。规则、似菌丝粉 17.不规则、卷发状、边缘波状18。不规则、呈丝状19。不规 臀“国养 则、根 一定培养条 下形成 此可观 南构形态荧边 等。 菌落的特征对菌 鉴定有 不同细茵种类的茵落各不相同,同 一种细茵因培养基的成分和 表面湿度的不同,菌落形态也有变化。同一种细菌在同一培养基上形成的菌落一般表现为相同的茵落形态特征,是鉴 定菌种的形态标志之 例如园裙固每菌(420tob2 cter chr0oe0Ccu题)在阿须贝氏于每培装基上表面茵落呈盐理热 状,凸起,边缘整齐,表面起初为光滑无色,以后逐渐产生皱折和产生黑色素。又例如蜡质芽孢杆菌霉状变种(Bacillus cereus var,,ycoides)在牛肉膏蛋白陈培养基表面形成类似菌丝体的菌落,从中央向四周弯曲伸延,铺展在培养基 表面上。有些细南产生色素。有些色素是水不溶性的,存在于菌体内如光合细菌的光合色素,赛氏杆菌(Serratia 有 2素是水溶性 扩散到培养 单胞菌(Pseudomonas chlororaphis)分 单胞菌 R.fluorescens) 分荧光色素到养中 】 细菌的液 因菌种及需氧性等去出不同的 。当菌体大量增殖时 有的形成均 油液 有的形成 在西生长时还的讲液生有跑形 的形成画浮 生液体表面 、碱和色素等。图1.24细菌在肉汤培养基中的生长 第二节放线幽 放线菌(Actinomycete)菌体形态为分枝的丝状体,属于原核微生物。放线菌革兰氏染 色都呈阳性反应,不运动,大部分是腐生菌,少数为寄生菌。放线菌对国民经济的重要性, 繁状 环状 浮膜状 在于它们是抗生素的主要产生菌,许多在临床和农业生产上有使用价值的抗生素都是由放线 在番体转化、石油脱蜡、烃类发酵、污水处 方面也有所应用 固定 由于放线 等方面有者重要的作 的 蜡、琼脂 复有机物 此外 少数放线菌也能引起人 畜和植物 放线菌的菌体为单细胞,最简单的为杆状或有原始菌丝,大部分放线茵由分枝发达的菌丝组成。茵丝无隔膜,菌 丝直径与杆状细菌差不多,大约】μm 细胞壁中含有N乙酰胞壁酸与二氨基庚二酸,而不含几丁质与纤维素 链霉菌属(Streptomyces)是放线菌中发有较为高等的放线菌,这里以其为例来阐明放线菌的一般形态构造。根据 放线菌茵丝的形态与功能不同,分为基内菌丝、气生菌丝与孢子丝。 (一)、基内菌丝 基内菌丝(substrate mycelium)又称营养菌丝(vegetative mycelium)或初级菌丝(primary mycelium),生长于
球菌等。 少数种类如柄细菌分裂后产生一个有柄不运动和一个无柄有鞭毛的子细胞,称为异形分裂。此外还有通过出芽方 式进行繁殖,如芽生杆菌 ( Blastobacter ) 、生丝微菌 ( Hyphomicrobium ) 的芽殖,蛭弧菌侵入宿主细菌细胞的 壁与膜间隙生长、分裂、产生多个子细胞的多次分裂以及节杆菌 ( Arthrobacter ) 的劈裂 (Snapping division) 等 特殊的繁殖方式。 细菌除无性繁殖外,电镜观察和遗传学研究已证明细菌存在着有性接 合。然而细菌有性接合较少,以无性繁殖为主。 图 1.22 细菌细胞的裂殖, ↑ 示正在分裂, ↑↑ 示已经分裂。 ( 引自谢念铭主编《医学细菌电镜图谱》,人民卫生出版社, 1994) (二)、细菌的群体特征 1 、细菌菌落特征 细菌在固体培养基上生长发育,几天内即可由一个或几个细菌分裂繁 殖成千上万个细胞,聚集在一起形成肉眼可见的群体,称为菌落 (colony) 。如果一个菌落是由一个细菌菌体生长、 繁殖而成,则称为纯培养。因此,可以通过单菌落计数的方法来计数细菌的数量。在微生物的纯种分离中也可以挑起 单个菌落进行移植的方法来获得纯培养物。 图 1 · 23 细菌菌落特征 正面观: 1. 扁平 2. 隆起 3. 低凸起 4. 高凸起 5. 脐状 6. 草 帽状 7. 乳头状 表面结构、形态及边缘: 8. 圆形、边缘完整 9. 不规则、边缘波 浪 10. 不规则、 颗粒状 、边缘叶状 11. 规划、放射状、边缘呈 叶状 12. 规则、边缘呈扇边状 13. 规则、边缘呈齿状 14. 规则、 有同心环、边缘完整 15. 不规则、似毛毯状 16. 规则、似菌丝状 17. 不 规则、卷发状、边缘波状 18. 不规则、呈丝状 19. 不规 则、根状 各种细菌在一定培养条件下形成的菌落具有一定的特征 ( 图 1.23) ,包括菌落的大小、 形状、光泽、颜色、硬度、透明度等 等。菌落的特征对菌种识别、鉴定有一定意义。 不同细菌种类的菌落各不相同,同一种细菌因培养基的成分和 表面湿度的不同,菌落形态也有变化。同一种细菌在同一培养基上形成的菌落一般表现为相同的菌落形态特征,是鉴 定菌种的形态标志之一。例如园褐固氮菌 ( Azotobacter chroococcum ) 在阿须贝氏无氮培养基上表面菌落呈粘稠糊 状,凸起,边缘整齐,表面起初为光滑无色,以后逐渐产生皱折和产生黑色素。又例如蜡质芽孢杆菌霉状变种 ( Bacillus cereus var, mycoides ) 在牛肉膏蛋白胨培养基表面形成类似菌丝体的菌落,从中央向四周弯曲伸延,铺展在培养基 表面上。有些细菌产生色素。有些色素是水不溶性的,存在于菌体内如光合 细菌的光合色素,赛氏杆菌 ( Serratia marcescens ) 的灵菌素;有些色素是水溶性,扩散到培养基中,例如绿色假单胞菌 ( Pseudomonas chlororaphis ) 分 泌绿色素到培养基中,荧光假单胞菌 ( P.fluorescens ) 分泌荧光色素到培养基中。 2 、细菌的液体培养特征 细菌在液体培养基中生长,因菌种及需氧性等表现出不同的特征。当菌体大量增殖时, 有的形成均匀一致的混浊液;有的形成沉淀;有的形成菌膜漂浮在液体表面( 图 1.24) 。有 些细菌在生长时还可同时产生气泡、酸、碱和色素等。图 1.24 细菌在肉汤培养基中的生长 第二节 放线菌 放线菌 (Actinomycete) 菌体形态为分枝的丝状体,属于原核微生物。放线菌革兰氏染 色都呈阳性反应,不运动,大部分是腐生菌,少数为寄生菌。放线菌对国民经济的重要性, 在于它们是抗生素的主要产生菌,许多在临床和农业生产上有使用价值的抗生素都是由放线 菌产生的。放线菌还可用于生产各种酶和维生素,在甾体转化、石油脱蜡、烃类发酵、污水处理等方面也有所应用。 有的菌还能与植物共生,固定大气氮。由于放线菌有很强的分解纤维素、石蜡、琼脂、角蛋白和橡胶等复杂有机物的 能力,故它们在自然界物质循环和提高土壤肥力等方面有着重要的作用。此外,少数放线菌也能引起人,畜和植物疾 病,如马铃薯疮痂病和人畜共患的诺卡氏菌病等。 一、放线菌的形态构造 放线菌的菌体为单细胞,最简单的为杆状或有原始菌丝,大部分放线菌由分枝发达的菌丝组成。菌丝无隔膜,菌 丝直径与杆状细菌差不多,大约 1 μ m 。细胞壁中含有 N- 乙酰胞壁酸与二氨基庚二酸,而不含几丁质与纤维素。 链霉菌属 ( Streptomyces ) 是放线菌中发育较为高等的放线菌,这里以其为例来阐明放线菌的一般形态构造。根据 放线菌菌丝的形态与功能不同,分为基内菌丝、气生菌丝与孢子丝。 (一) 、基内菌丝 基内菌丝(substrate mycelium)又称营养菌丝(vegetative mycelium)或初级菌丝 (primary mycelium),生长于 培养基内,主要功能为吸收营养物。链霉菌基内菌丝一般无隔膜,多分枝,直径常在 0.2~1.0 μm 。有的无色,有的 能产生色素,呈红、橙、黄、绿、蓝、紫、褐、黑等不同颜色。色素有水溶性的,也有脂溶性的。若是水溶性的色素

则可渗入培养基内,将培养基染上相应的颜色:如是非水溶性的(或脂溶性)色素,则使菌落呈现相应的颜色。不 同类型的放线菌基内菌丝的形态特征有所区别,例如诺卡氏(Vocardia)基内菌丝强烈弯曲如树根状,生长到 ,又如束丝放线菌(ctinosynnema)基丝可参与气丝一起扭成菌 色素。各类放线菌能否产生菌丝体,取决于种的特征,营养条件和环境因子 (三)、孢子丝 放线南生长至一定阶段,在其气生菌丝上分化出可以形成孢子的菌丝,为孢子丝。孢子丝的形状以及在气生南丝上的 排列方式,随不同菌种而不同。孢子丝的形状有直形、波浪形、螺旋形之分(图1.25)。螺旋状孢子丝的螺旋结构 与长度均很稳定,螺旋数目、疏密程度、旋转方向等都是种的特征。孢子丝的排列方式,有的交替着生,有的丛生或 点分出3个以上的孢子枝者,称轮生枝。它有一级轮生和二级轮生之分。轮生类群的孢子丝多为 定阶段断裂为孢子 孢子有球形、 圆形、杆形、瓜子形等不同形状。在电子显 微镜下可见孢子表面结构,有的光滑、有的 带小疣、有的生刺或毛发状。孢子常具有不 同色素。孢子形状、表面结构、颜色等均为 鉴定放线菌菌种的依据。 二、放线菌的繁殖与菌落特征 放线菌主要通过无性孢子及菌丝片段进 轮生,无 孢子在 繁旋 丝分裂成为 中吸收水 14根芽管 。少数放线黄 先在菌丝上形成孢子囊,在孢子囊内形成孢 囊孢子。孢子囊可在气生菌丝上形成,也可 在营养茵丝上形成,或一者均可生成。狗 囊成熟后,释放出大量孢囊孢子。孢囊孢子 可萌发形成菌丝体 螺单轮 图1.26链霉的生活史简图 无螺的二级轮 内菌丝体3 气生菌丝体 孢子正 孢子丝分化为子形成 。 T形成 因而液体发酵就是利用 方式进行增殖的 放线菌菌落在光学显微镜下观察,周围具放射状菌丝。放线南菌落因种类不同可分 为两 类是由产生大量分枝的气生菌丝的茵种所形成的菌落,以链霉菌的菌落为代 表。链霉菌菊丝较细,生长缓慢,南丝分枝互相交错缠绕,因而形成的南落质地致:, 表面呈紧密的绒状或坚实、干燥、多皱,菌落较小而不致广泛延伸:营养菌丝长在培养 所以菌落与培养基结合较紧,不易挑起或整个茵落被挑起而不致破碎。幼令菌 类的孢子含有色素 使形成 粉末状或粒状的奥 的菌落,因其一般只有基内菌丝, 粘若力差,结构呈粉质状, 针排起则易粉 三、放线菌的主要类群 链幕菌属(Strepto 链霉菌属有发有良好的分枝状菌丝体,菌丝无隔膜,直径约0.4-1μm ,长短不一,多核。菌丝体有营养菌丝 气生菌丝和孢子丝之分。孢子丝再形成分生孢子。链莓菌主要借分生孢子繁殖,其生活史见图1·6
则可渗入培养基内,将培养基染上相应的颜色;如是非水溶性的 ( 或脂溶性 ) 色素,则使菌落呈现相应的颜色。不 同类型的放线菌基内菌丝的形态特征有所区别,例如诺卡氏菌 ( Nocardia ) 基内菌丝强烈弯曲如树根状,生长到一 定菌令后,产生横隔膜,并断裂成不同形状的杆菌体。又如束丝放线菌(Actinosynnema ) 基丝可参与气丝一起扭成菌 丝束,屹立在基质表面,好似刚出土的“竹笋”状等等。 (二) 、气生菌丝 气生菌丝 (aerial mycelium) 又称二级菌丝 (secondary mycelium) 。由基内菌丝长出培养基 外伸向空间的菌 丝为气生菌丝。在显微镜下观察时,气生菌丝体颜色较深,直径较基内菌丝粗,约 1~1.4 μm,直或弯曲,有的产生 色素。各类放线菌能否产生菌丝体,取决于种的特征,营养条件和环境因子。 (三) 、孢子丝 放线菌生长至一定阶段,在其气生菌丝上分化出可以形成孢子的菌丝,为孢子丝。孢子丝的形状以及在气生菌丝上的 排列方式,随不同菌种而不同。孢子丝的形状有直形、波浪形、螺旋形之分 ( 图 1.25) 。螺旋状孢子丝的螺旋结构 与长度均很稳定,螺旋数目、疏密程度、旋 转方向等都是种的特征。孢子丝的排列方式,有的交替着生,有的丛生或 轮生。孢子丝从一点分出 3 个以上的孢子枝者,称轮生枝。它有一级轮生和二级轮生之分。轮生类群的孢子丝多为二 级轮生。这些特征,均为放线菌菌种鉴定的依据。 1.25 放线菌孢子丝的类型 孢子丝生长到一定阶段断裂为孢子,或 称分生孢子 (conidium) 。孢子有球形、椭 圆形、杆形、瓜子形等不同形状。在电子显 微镜下可见孢子表面结构,有的光滑、有的 带小疣、有的生刺或毛发状。孢子常具有不 同色素。孢子形状、表面结构、颜色等均为 鉴定放线菌菌种的依据。 二、放线菌的繁殖与菌落特征 放线菌主要通过无性孢子及菌丝片段进 行繁殖。用电子显微技术和超薄切片的研究 表明,放线菌通过产生横隔膜的方式使孢子 丝分裂成为一串分生孢子。孢子在适宜环境 中吸收水分,膨胀萌发,长出 1~4 根芽管, 形成新的菌丝体 ( 图 1.26)。少数放线菌首 先在菌丝上形成孢子囊,在孢子囊内形成孢 囊孢子。孢子囊可在气生菌丝上形成,也可 在营养菌丝上形成,或二者均可生成。孢子 囊成熟后,释放出大量孢囊孢子。孢囊孢子 可萌发形成菌丝体。 图 1.26 链霉菌的生活史简图 1. 孢子萌发 2. 基内菌丝体 3. 气生菌丝体 4. 孢子丝 5. 孢子丝分化为孢子 初级的放线菌如放线菌属 (Actinomyces) 、分枝杆菌属 (Mycobacterium) 只形成 短小分枝或基内菌丝并通过细胞分裂或菌丝断裂来繁殖。放线菌的菌丝片段可形成新的 菌丝体。在液体振荡培养成工业发酵时很少形成分生孢子,因而液体发酵就是利用这一 方式进行增殖的。 放线菌菌落在光学显微镜下观察,周围具放射状菌丝。放线菌菌落因种类不同可分 为两类。一类是由产生大量分枝的气生菌丝的菌种所形成的菌落,以链霉菌的菌落为代 表。链霉菌菌丝较细,生长缓慢,菌丝分枝互相交错缠绕,因而形成的菌落质地致密, 表面呈紧密的绒状或坚实、干燥、多皱,菌落较小而不致广泛延伸;营养菌丝长在培养 基内,所以菌落与培养基结合较紧,不易挑起或整个菌落被挑起而不致破碎。幼令菌落 因气生菌丝尚未分化成孢子丝,故菌落表面与细菌菌落相似而不易区分。当形成大量孢 子布满菌落表面时,就形成外观为绒状、粉末状或颗粒状的典型的放线菌菌落;有些种 类的孢子含有色素,如与基内菌丝的颜色不同,则使菌落表面与背面呈现不同颜色。另一类菌落由不产生大量菌丝体 的种类形成,如诺卡氏菌的菌落,因其一般只有基内菌丝,结构松散,粘着力差,结构呈粉质状,用针挑起则易粉碎。 放线菌菌落常具土腥味。 三、放线菌的主要类群 (一) 、链霉菌属 (Streptomyces) 链霉菌属有发育良好的分枝状菌丝体,菌丝无隔膜,直径约 0.4-1μm ,长短不一,多核。菌丝体有营养菌丝、 气生菌丝和孢子丝之分。孢子丝再形成分生孢子。链霉菌主要借分生孢子繁殖,其生活史见图 1·26

各种碳刻 多者 每系 小单孢黄属c2m ora) 该其内丝发有自好多公 无墙隔。不 断裂,直径为03 般不形成气生菌丝体 孢子单生,无柄,直接从基内菌丝上产生,或在基内 茵丝上长出短孢子梗,顶端着生一个孢子(图127) 小单孢菌属与链霉菌属相比,菌丝体较细、无气 生菌丝:菌落小,一般为23mm,呈橙黄色或红色, 也有深褐、黑色、蓝色者:菌丝生长力较弱, 一般在 520 天便停 ,生长温度略高, 一般为32℃ 37℃,所得布于士或堆肥中。庆大霉素即由棘孢 图128诺卡氏菌的形 图1.27小单孢南的形态 小单孢菌 生 t al 2000 (三)、诺卡氏黄属(ocardia) 诺卡氏菌在培养基上形成典型的黄丝体,南丝纤细,多数弯曲如树根状,生长到十几小时时开始形成横隔膜,并 断裂成多形态的杆状、球状或带叉的杆状体。诺卡氏茵属中大多数种无气生菌丝,只有基内菌丝,菌落秃裸:有的则 在基内菌丝体上覆盖着极薄一层气生菌丝,有横隔,断裂成杆状(图1.28)。菌落比链霉菌的小,表面多皱,致密 干燥,或平滑凸起不等,有黄、黄绿、红橙等颜色 利福莓素由地中海诺卡氏菌(化mediterranei) 产生。有些诺卡氏菌可用于石油脱蜡、烃类发酵以及污水处理中 分 化合移 放线 属(ct 在 一般为厌氧或兼性厌氧菌,因此 放线茵属多为致病菌 原始发印 的而肿病」 个是衣氏放线菌(4C ,寄生在人体上,可引起后颚骨瘤肿病和肺脏 及胸部的放线菌病。 图1.29游动放线菌属的形态(引自Prescott et al..,2002 (五)、游动放线菌属(Actinoplanes) A.游动放线菌孢囊的发有B.游动放线菌孢囊孢子成熟并释放出来 游动放线菌属以基内菌丝为主,有的有气生菌丝,有的气 生菌丝少,菌丝有隔或无隔。在基内菌丝上生孢囊梗,梗顶端 生孢囊,孢囊成熟,释放出有鞭毛、在水中能运动的游动孢子, 见图1.29. 第三节古 现用 )这一概 今是沃断(W 类群的16SrRN 碱基序列进行研究比拉后 人认为。生物的发有不是 个简单的由原核生物发有到更 B 完全更复杂的真核生物的过程,而是明显地存在三个发育不同的基因系统:细菌、古南和真核生物。从发育的观点看 这三个举型中任何 类都不出其他两类出现得更古老。 一、古菌的一般特性和分类 古菌与细菌和真核微生物之间的异同已在本章前面详细阐述。可见古茵是一群具有独特基因结构或系统发育生物 的官松的量跑生物,大多生活在地球上如超高温、高酸藏度、高盐浓度、严格无氧状态等极环境或生命出现
已知的链霉菌属的菌有千余种,大多生长在含水量较低、通气良好的土壤中。链霉菌能分解纤维素、石蜡、蜡与 各种碳氢化合物。链霉菌是产生抗生素菌株的主要来源。许多著名的常用的抗生素如链霉素、土霉素,抗肿瘤的博来 霉素、丝裂霉素,抗真菌的制霉菌素,抗结核的卡那霉素,能有效防治水稻纹枯病的井冈霉素等,都是链霉菌属的种 的次生代谢产物。 (二) 、小单孢菌属 (Micromonospora) 该属菌基内菌丝发育良好,多分枝,无横隔,不 断裂,直径为 0.3~0.6μm,一般不形成气生菌丝体。 孢子单生,无柄,直接从基内菌丝上产生,或在基内 菌丝上长出短孢子梗,顶端着生一个孢子(图 1.27) 。 小单孢菌属与链霉菌属相比,菌丝体较细、无气 生菌丝;菌落小,一般为 2~3mm ,呈橙黄色或红色, 也有深褐、黑色、蓝色者;菌丝生长力较弱,一般在 15~20 天便停止发育,生长温度略高,一般为 32℃~ 37℃,所以很易区别开。 此属多分布于土壤或堆肥中。庆大霉素即由棘孢 小单孢菌 (Micromonospora echinospora)产生。 (三) 、诺卡氏菌属(Nocardia) 诺卡氏菌在培养基上形成典型的菌丝体,菌丝纤细,多数弯曲如树根状,生长到十几小时时开始形成横隔膜,并 断裂成多形态的杆状、球状或带叉的杆状体。诺卡氏菌属中大多数种无气生菌丝,只有基内菌丝,菌落秃裸;有的则 在基内菌丝体上覆盖着极薄一层气生菌丝,有横隔,断裂成杆状 ( 图 1.28) 。菌落比链霉菌的小,表面多皱,致密 干燥,或平滑凸起不等,有黄、黄绿、红橙等颜色。 利福霉素由地中海诺卡氏菌 (N.mediterranei) 产生。有些诺卡氏菌可用于石油脱蜡、烃类发酵以及污水处理中 分解腈类化合物。 (四) 、放线菌属(Actinomyces) 该属仅有基内菌丝,有横隔,易断裂成“ V ”形或“Y”形体。菌落污白色。一般为厌氧或兼性厌氧菌,因此, 在 CO2 气体存在下容易生长。放线菌属多为致病菌。典型种为牛型放线菌 ( Actinomyces bovis ) ,原始发现于牛 的颚肿病,通常见于动物口腔内。另一个是衣氏放线菌 (Act.israeli),寄生在人体上,可引起后颚骨瘤肿病和肺脏 及胸部的放线菌病。 图 1.29 游动放线菌属的形态 ( 引自 Prescott et al., 2002) (五) 、游动放线菌属 (Actinoplanes) A. 游动放线菌孢囊的发育 B. 游动放线菌孢囊孢子成熟并释放出来 游动放线菌属以基内菌丝为主,有的有气生菌丝,有的气 生菌丝少,菌丝有隔或无隔。在基内菌丝上生孢囊梗,梗顶端 生孢囊,孢囊成熟,释放出有鞭毛、在水中能运动的游动孢子, 见图 1.29 。 第三节 古菌 “古细菌”(archaebacteria ,现用 archaea)这一概 念是沃斯(Woese)及他的同事们对代表性细菌类群的 16S rRNA 碱基序列进行研究比较后于 1977 年提出来的。沃斯(Woese) 等人认为,生物界的发育不是一个简单的由原核生物发育到更 完全更复杂的真核生物的过程,而是明显地存在三个发育不同的基因系统:细菌、古菌和真核生物。从发育的观点看, 这三个类型中任何一类都不比其他两类出现得更古老。 一、古菌的一般特性和分类 古菌与细菌和真核微生物之间的异同已在本章前面详细阐述。可见古菌是一群具有独特基因结构或系统发育生物 大分子序列的单细胞生物,大多生活在地球上如超高温、高酸碱度、高盐浓度、严格无氧状态等极端环境或生命出现 初期的自然环境。 图 1.27 小单孢菌的形态 图 1.28 诺卡氏菌的形态 ( Madigan et al., 2000) A. B

古菌是一大类形态各异、特殊生理功能绝然不同的微生物群,如产甲烷细菌,可在严格厌氧环境下利用简单二碳 ~碳化合物或C02生存和产甲烷:还原硫酸盐古菌可在极端高温、酸性条件下还原硫酸盐:极端嗜盐古菌可在极高 存,等。古可营自养或异养型生 嗜盐杆菌黑 胞壁 细胞草 而致细的多彩 使有 盐球菌 能是由 有细单位组成或由假胞壁质组 细韵的化学组成上 吉细菌含有异戊烯而不含脂肪酸 甲统杆 硫化叶菌调 酯,脂肪酸也为有分支的直链而不是无分支的直链。16S rRNA的碱基序列, tNA的特殊其的修饰。5 S TRNA的 二级结构等均不同干细菌和点核尚牛物。对干各种抗生表 的敏感性上也与细菌有很大差异,如对于氯莓素、青霉素、 利福平等抗生素不敏感,但细菌对此敏感:相反对于环已 热原体 还原球菌 胺、苗香莓素等敏感而细菌却不敏感 根据不园的生特性 可将 为5 大类群: 烷 古生古界 菌群和极滞热和 热代谢元素硫 图1.30古菌的系统发育树(Madigan et al.,2000) 立国检士细曲联 产甲烷菌是一群迄今为止所知的最严格厌氧的、能形成甲烷的化能自养或化能异养的古菌群 一)、产甲烷细菌的形态与细胞结构 产甲烷细菌是 一大都能产生甲烷的生理类群,因此包括了球形、杆形、螺旋形、长丝状等不同形态,见图1.31。 产甲烷细菌的细胞壁染色后有围绕细胞约有15n一20m厚的一层无定形电子稠密层。在化学组成上含有假胞壁 质。假胞壁质与细菌的胞壁质在化学结构上有区别:假胞壁质肽链上为L-氨基酸,糖链上为 乙酰L塔罗糖形 糖醛酸,而且连接此N 乙酰 乙酰-葡萄糖胺的糖苷键主要为B1,3)糖苷键,少部 分为糖 糖链上为 乙酰胞 外膜 执 )的细胞壁为均匀的单层,厚度为 氏甲 壁具有2个电子致密层和1个电子透明的中层。瘤胃甲烷短杆菌细胞壁由电子致密层的内层,电子透明层的中层和 粗糙不规则形的外层构成,索氏甲烷丝菌细胞壁由电子密度较高并与质膜相邻的内层和电子密度较低的外层构成。亨 氏甲烷螺菌(旅thanospirillum hungatii)有内壁和外壁之分,外壁在菌丝体处连续成鞘状,不重叠凹陷,内壁仅 围绕丝状体中的各个细胞,细胞横隔处只见外壁。在不同产甲烷细菌细胞壁表层还有不同的物质层包围。绝大多数甲 翻居修球使不田业中塞教有素成的松第拜列的多聚合所包通,业中塞育 乙酰半乳湛胺、D一面 菌有 白表层包 有特别高的 o在aac)特甲烧菌由具有韩异嘴 的鞘包, 甘 产甲烷细菌和其他古细 群的细胞原化学组成与其他生物细胞膜明显不同。 古菌膜脂的丰要成分为植烷醇基甘油 醚,包括C20的二植烷基甘油醚和C40的双二植烷基二甘油四醚。这种脂类缺乏可皂化的脂,羟基被甘油磷酸醚 化,另一个经糖苷健和1个二糖相连。这样形成的磷酸甘油酯可以增进膜的刚性和稳定性。而其他生物质膜的主要成 分是款脂酸甘油酯
古菌是一大类形态各异、特殊生理功能绝然不同的微生物群,如产甲烷细菌,可在严格厌氧环境下利用简单二碳 和一碳化合物或 CO2 生存和产甲烷;还原硫酸盐古菌可在极端高温、酸性条件下还原硫酸盐;极端嗜盐古菌可在极高 盐浓度下生存,等等。古菌可营自养或异养型生活。古菌 具有独特的细胞或亚细胞结构,如无细胞壁古细菌没有细 胞壁,仅有细胞膜,而致细胞多形态。即使有细胞壁,可 能是由蛋白质亚单位组成或由假胞壁质组成,无胞壁酸。 细胞膜的化学组成上,古细菌含有异戊烯醚而不含脂肪酸 酯,脂肪酸也为有分支的直链而不是无分支的直链。16S rRNA 的碱基序列, tRNA 的特殊碱基的修饰, 5S rRNA 的 二级结构等均不同于细菌和真核微生物。对于各种抗生素 的敏感性上也与细菌有很大差异,如对于氯霉素、青霉素、 利福平等抗生素不敏感,但细菌对此敏感;相反对于环己 胺、茴香霉素等敏感而细菌却不敏感。 根据不同的生理特性,可将古菌分为 5 大类群:产甲 烷古菌群、还原硫酸盐古菌群、极端嗜盐古菌群、无细胞 壁古菌群和极端嗜热和超嗜热代谢元素硫古菌群。它们之 间的系统发育关系如图 1.30 所示。 图 1.30 古菌的系统发育树 ( Madigan et al., 2000) 二、产甲烷古细菌群 产甲烷菌是一群迄今为止所知的最严格厌氧的、能形成甲烷的化能自养或化能异养的古菌群。 (一)、产甲烷细菌的形态与细胞结构 产甲烷细菌是一大都能产生甲烷的生理类群,因此包括了球形、杆形、螺旋形、长丝状等不同形态,见图 1.31 。 产甲烷细菌的细胞壁染色后有围绕细胞约有 15nm-20nm 厚的一层无定形电子稠密层。在化学组成上含有假胞壁 质。假胞壁质与细菌的胞壁质在化学结构上有区别:假胞壁质肽链上为 L- 氨基酸,糖链上为 N- 乙酰 -L- 塔罗糖胺 糖醛酸,而且连接此 N- 乙酰 -L- 塔罗糖胺糖醛酸和 N- 乙酰 -D- 葡萄糖胺的糖苷键主要为 β1,3)糖苷键,少部 分为 β1,4)糖苷键。细菌胞壁质肽链上为 D- 氨基酸,糖链上为 N- 乙酰胞壁酸,连接的糖苷键与产甲烷细菌的相 同。产甲烷细菌无细菌细胞壁胞壁质特征而富含各种表层蛋白。产甲烷细菌的外膜可分为 5 种类型:①固定结构层 + 细胞质膜;②亚甲基软骨素层 + S 层 + 细胞质膜;③固定结构层 + 假胞壁质 + 细胞质膜;④鞘 +S 层 + 细胞质膜 和⑤假胞壁质 + 细胞质膜。嗜热自养甲烷杆菌( Methanobacterium thermoautotrophicum )和布氏甲烷短杆菌( Mb. bryantii )的细胞壁为均匀的单层,厚度为 10nm ~18nm 。史氏甲烷短杆菌( Methanobrevibacter smithii )细胞 壁具有 2 个电子致密层和 1 个电子透明的中层。瘤胃甲烷短杆菌细胞壁由电子致密层的内层,电子透明层的中层和 粗糙不规则形的外层构成,索氏甲烷丝菌细胞壁由电子密度较高并与质膜相邻的内层和电子密度较低的外层构成。亨 氏甲烷螺菌( Methanospirillum hungatii )有内壁和外壁之分,外壁在菌丝体处连续成鞘状,不重叠凹陷,内壁仅 围绕丝状体中的各个细胞,细胞横隔处只见外壁。在不同产甲烷细菌细胞壁表层还有不同的物质层包围。绝大多数甲 烷八叠球菌被一个由亚甲基软骨素组成的松紧排列的多聚合网所包围,亚甲基软骨素是由 N- 乙酰半乳糖胺、 D- 葡 糖醛酸(或 D- 半乳糖醛酸)、微量 D- 葡萄糖、痕量 D- 甘露糖组成的无硫聚合物。甲烷嗜热菌属( Methanothermus ) 的 2 株古菌有一层糖蛋白表层包围,在这些糖蛋白中含有特别高的异亮氨酸、天冬氨酸和半胱氨酸,其二级结构中仅 含 7% 的 α - 螺旋,而 β - 螺旋却高达 44% ,这有利于在极端嗜热条件下的生存。不同产甲烷细菌的细胞壁构成 见表 1-6 。亨氏甲烷螺菌和索琴氏甲烷丝菌( Methanothrix soehngenii )等产甲烷细菌由具有表面特异超微结构 的鞘包裹,每个鞘内有多个甚至几十个连成丝状的细胞。 产甲烷细菌和其他古细菌群的细胞膜化学组成与其他生物细胞膜明显不同。古菌膜脂的主要成分为植烷醇基甘油 醚,包括 C 20 的二植烷基甘油醚和 C 40 的双二植烷基二甘油四醚。这种脂类缺乏可皂化的脂,羟基被甘油磷酸醚 化,另一个经糖苷键和 1 个二糖相连。这样形成的磷酸甘油酯可以增进膜的刚性和稳定性。而其他生物质膜的主要成 分是软脂酸甘油酯
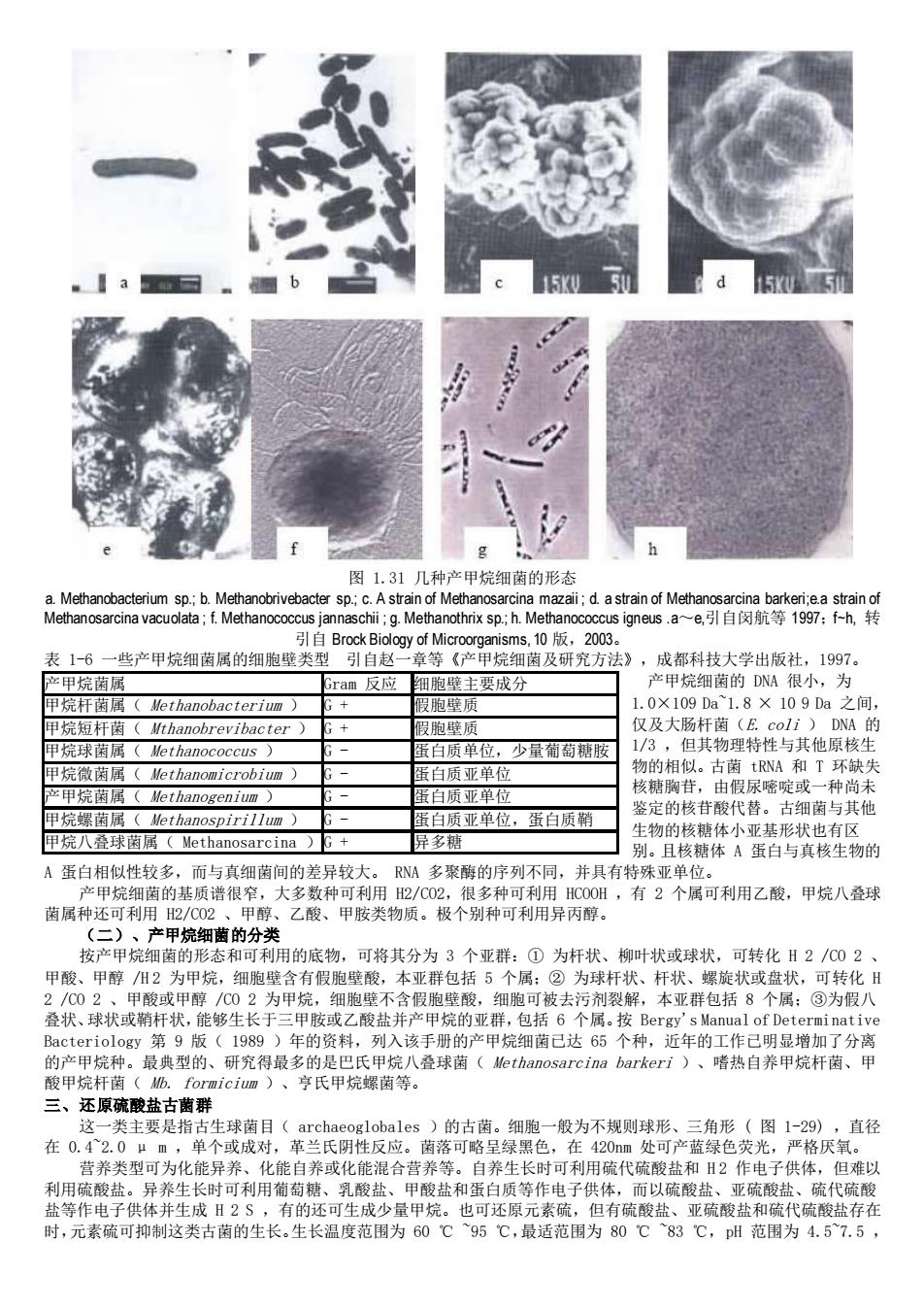
15U50 15KU5 图1.31几种产甲烷细菌的形态 .9 thnx sp. 表1-6一些产甲烷细属的细胞壁类型 白 等产里烷细菌及研究方法》 成都科技大学出版补,1997 ram反应 胞壁主要成分 产甲烷细菌的DNA很小,为 甲烷杆菌 1.0×109Da1.8×109Da之间, 仅及大肠杆菌(E.coli)DNA的 质单位,少量葡萄糖服 1/3,但其物理特性与其他原核生 甲烷微菌属(ethano咖icrobium) 物的相似。古菌tRNA和T环缺失 甲烷螺菌属(Methanospirillum) 白质亚单位,蛋白质箱 甲烷八叠球菌属(Methanosarcina)G+ 异多糖 物的个蛋资的 A蛋白相似性较多,而与真细菌间的差异较大。 甲棕细的基质造 RNM多聚酶的序列不同,并具有特殊亚单位 大多数种 有2个属可利用乙酸,甲烷八叠球 茵属种还可利用2/C02、 乙酸、甲胺类物质。极个别种可利用异丙醇。 (二)、产甲烷细菌的分类 按产甲烷细菌的形态和可利用的底物,可将其分为3个亚群:①为杆状、柳叶状或球状,可转化H2/C02、 甲酸、甲醇州2为甲烷,细胞壁含有假胞壁酸,本亚群包括5个属:②为球杆状、杆状、螺旋状或盘状,可转化日 2/002、甲酸或甲醇/02为甲烷,细胞壁不含假胞壁酸,细胞可被去污剂裂解,本亚群包括8个属:③为假八 叠状、球状或鞘杆状,能够生长于 乙酸盐并产甲烷的亚群,包括6个属.按Bergy's Manual of Determinative 年的资料列入该手册的产甲烷细己达65个种,近年的工作己明显增加了分离 阳C用综种。最典型的、研究得最多的是里菌等。 八叠球菌(Hethanosarcina barkeri)、嗜热自养甲烷杆菌、甲 这一类主要是 一股为不规则球形 直径 在0.42.0um 单个或成对,革兰氏阴性反应。陶落可略呈绿黑色,在42Om处可产蓝绿色荧光,严格厌 营养类型可为化能异养、化能自养或化能混合营养等。自养生长时可利用硫代硫酸盐和Ⅱ2作电子供体,但难以 利用硫酸盐。异养生长时可利用葡萄糖、乳酸盐、甲酸盐和蛋白质等作电子供体,而以硫酸盐、亚硫酸盐、硫代硫酸 盐等作电子供体并生成H2S,有的还可生成少量甲烷。也可还原元素硫,但有硫酸盐、亚硫酸盐和硫代硫酸盐存在 时,元素疏可抑制这类古菌的生长。生长温度范围为60℃95℃,最适范围为80℃83℃,DH范围为4.57.5
图 1.31 几种产甲烷细菌的形态 a. Methanobacterium sp.; b. Methanobrivebacter sp.; c. A strain of Methanosarcina mazaii ; d. a strain of Methanosarcina barkeri;e.a strain of Methanosarcina vacuolata ; f. Methanococcus jannaschii ; g. Methanothrix sp.; h. Methanococcus igneus .a~e,引自闵航等 1997;f~h, 转 引自 Brock Biology of Microorganisms, 10 版,2003。 表 1-6 一些产甲烷细菌属的细胞壁类型 引自赵一章等《产甲烷细菌及研究方法》,成都科技大学出版社,1997。 产甲烷细菌的 DNA 很小,为 1.0×109 Da~1.8 × 10 9 Da 之间, 仅及大肠杆菌(E. coli ) DNA 的 1/3 ,但其物理特性与其他原核生 物的相似。古菌 tRNA 和 T 环缺失 核糖胸苷,由假尿嘧啶或一种尚未 鉴定的核苷酸代替。古细菌与其他 生物的核糖体小亚基形状也有区 别。且核糖体 A 蛋白与真核生物的 A 蛋白相似性较多,而与真细菌间的差异较大。 RNA 多聚酶的序列不同,并具有特殊亚单位。 产甲烷细菌的基质谱很窄,大多数种可利用 H2/CO2,很多种可利用 HCOOH ,有 2 个属可利用乙酸,甲烷八叠球 菌属种还可利用 H2/CO2 、甲醇、乙酸、甲胺类物质。极个别种可利用异丙醇。 (二)、产甲烷细菌的分类 按产甲烷细菌的形态和可利用的底物,可将其分为 3 个亚群:① 为杆状、柳叶状或球状,可转化 H 2 /CO 2 、 甲酸、甲醇 /H 2 为甲烷,细胞壁含有假胞壁酸,本亚群包括 5 个属;② 为球杆状、杆状、螺旋状或盘状,可转化 H 2 /CO 2 、甲酸或甲醇 /CO 2 为甲烷,细胞壁不含假胞壁酸,细胞可被去污剂裂解,本亚群包括 8 个属;③为假八 叠状、球状或鞘杆状,能够生长于三甲胺或乙酸盐并产甲烷的亚群,包括 6 个属。按 Bergy's Manual of Determinative Bacteriology 第 9 版( 1989 )年的资料,列入该手册的产甲烷细菌已达 65 个种,近年的工作已明显增加了分离 的产甲烷种。最典型的、研究得最多的是巴氏甲烷八叠球菌( Methanosarcina barkeri )、嗜热自养甲烷杆菌、甲 酸甲烷杆菌( Mb. formicium )、亨氏甲烷螺菌等。 三、还原硫酸盐古菌群 这一类主要是指古生球菌目( archaeoglobales )的古菌。细胞一般为不规则球形、三角形 ( 图 1-29) ,直径 在 0.4~2.0 μ m ,单个或成对,革兰氏阴性反应。菌落可略呈绿黑色,在 420nm 处可产蓝绿色荧光,严格厌氧。 营养类型可为化能异养、化能自养或化能混合营养等。自养生长时可利用硫代硫酸盐和 H 2 作电子供体,但难以 利用硫酸盐。异养生长时可利用葡萄糖、乳酸盐、甲酸盐和蛋白质等作电子供体,而以硫酸盐、亚硫酸盐、硫代硫酸 盐等作电子供体并生成 H 2 S ,有的还可生成少量甲烷。也可还原元素硫,但有硫酸盐、亚硫酸盐和硫代硫酸盐存在 时,元素硫可抑制这类古菌的生长。生长温度范围为 60 ℃ ~95 ℃,最适范围为 80 ℃ ~83 ℃,pH 范围为 4.5~7.5 , 产甲烷菌属 Gram 反应 细胞壁主要成分 甲烷杆菌属( Methanobacterium ) G + 假胞壁质 甲烷短杆菌( Mthanobrevibacter ) G + 假胞壁质 甲烷球菌属( Methanococcus ) G - 蛋白质单位,少量葡萄糖胺 甲烷微菌属( Methanomicrobium ) G - 蛋白质亚单位 产甲烷菌属( Methanogenium ) G - 蛋白质亚单位 甲烷螺菌属( Methanospirillum ) G - 蛋白质亚单位,蛋白质鞘 甲烷八叠球菌属( Methanosarcina )G + 异多糖