
构成分之外,许多蛋白质本身就是运输养料的透酶或具催化活性的酶蛋白,在细胞代谢过程中起者重要作用。 图L,l0细南细胞质膜的恭本结构(引自Prescott et al., 2002) 质和限的丰功能为:Q拉制细内、外的物 营养物 R 久 (I P 酸 电及化学物 他幸之生的清信是并发生执免李化细陶的化 代谢变化和产生相应的反应。 除细胞质膜外,很多细南还具有内膜系统。①中间体 (esosome)是由细胞膜局部内陷折叠而成,它与细胞壁的合成、 核质分裂、细胞呼吸以及芽孢形成有关。由于中间体具有类似真核细胞线粒体的作用,又称拟线粒体。②类囊体 (thylakoid)是蓝细菌细胞中存在的囊状体,由单位膜组成,上面分布有叶绿素、藻胆色素等光合色素和有关酶类 通透 磷酸 由蛋白质为主的单层 内含因固定2所围的5 酮糖激酶,是白养细南固定C02的场所 (三)细购质 细胞质(cytop1asm)是细胞膜内的物质,除细胞核外皆为细胞质。它无色透明,呈粘胶状,主要成分为水、蛋白 质、核酸、脂类,也含有少量的糖和盐类。由于富含核酸,因而嗜碱性强,幼令菌者色均匀。此外,细胞质内还含有 核糖体、颗粒状内含物和气泡等物质。 1、核糖体 核糖体(riboson©)亦称核蛋白体,为多肽和蛋白质合成的场所。在电子显微镜下可见到细菌的核糖体游离于细 分教 两亚单元组成化 体既可以游 状态存有 质中 也可结合于 质风 、内含 很多细古 在营养物质丰富的时候,其细胞内聚合各种不同的贮藏颗粒,当营养缺乏时,它们又能被分解利用。这 种贮藏颗粒可在光学显微镜下观察到,通称为内含物(cytoplasmic inclusions)。贮藏颗粒的多少可随菌令及培 养条件不同而改变。 异染颗粒 ,(metachromatic granules))又称捩转菌素(volutin),最早发现于迂回螺(Spirillum volutans) 中。异染粒是以无机偏磷酸盐聚合物为主要成分的一种无机磷的贮备物。异染颗粒咯碱性或嗜中性较强,用蓝色染料 (如甲苯胺蓝或甲烯蓝)染色后不呈蓝色而呈紫红色,故称异染颗粒 聚B-羟基丁酸 poly- wdro不若色 一种 和能 源性贮藏物。它是D3 苏丹黑着 细中的清 —羟基丁酸颗料 等细菌常积品见 1细碧内含物 Madigan et al. 2000)A 只能 在电镜下观察到,如用稀镇液染色成红揭色,可在光学显 感 微镜下看到。有的细南积累淀粉粒,用碘液可染成深蓝色。 聚B羟基丁酸期 肝糖粒、淀粉粒都是碳源贮藏物 流滴(sulfur globules) 某些氧化硫的细销细 胞内可积累硫滴。如贝氏硫菌属(Beggiatoa)、发硫 菌属(Thiothrix)在细胞内常含有强折光性的硫滴, 此为贮存的硫,系通过氧化硫化氢而形成,作为能量储备 细不体 存在王水生细利趋细黄 细菌 现出趋性、 。磁小体由 层含有碳脂、蛋白质和糖蛋白的膜包围」 3、气泡 某些水生细菌,如蓝细菌、不放氧的光合细菌和盐细菌细胞内贮存气体的特殊结构称气泡。气泡由许多小的的气 囊(gas vesicle)组成,气囊膜只含蛋白质而无碳脂。气泡的大小、形状和数量随细菌种类而异。气泡能使细胞保持 浮力,从而有助于调节并使细菌生活在它们需要的最佳水层位置,以利获得氧、光和营养。 (四)、细胞核 细菌细胞的核位于细胞质内,无核膜、无核仁,仅为一核区,因此称为原始形态的核(primitive form nucleus) 或拟核(ucleoid)。细菌细胞的原核只有一个染色体,主要含有具有遗传特征的脱氧核糖核酸(DA)。拟核中尚
构成分之外,许多蛋白质本身就是运输养料的透酶或具催化活性的酶蛋白,在细胞代谢过程中起着重要作用。 图 1.10 细菌细胞质膜的基本结构 ( 引自 Prescott et al., 2002) 细胞膜的主要功能为:① 控制细胞内、外的物质 ( 营养物 质和代谢废物 ) 的运送、交换;② 维持细胞内正常渗透压的屏 障作用;③ 合成细胞壁各种组分 (LPS 、肽聚糖、磷壁酸 ) 和 荚膜等大分子的场所;④ 进行氧化磷酸化或光合磷酸化的产能基 地;⑤ 传递信息。膜上的某些特殊蛋白质能接受光、电及化学物 质等产生的刺激信号并发生构象变化,从而引起细胞内的一系列 代谢变化和产生相应的反应。 除细胞质膜外,很多细菌还具有内膜系统。① 中间体 (mesosome) 是由细胞膜局部内陷折叠而成,它与细胞壁的合成、 核质分裂、细胞呼吸以及芽孢形成有关。由于中间体具有类似真核细胞线粒体的作用,又称拟线粒体。② 类囊体 (thylakoid) 是蓝细菌细胞中存在的囊状体,由单位膜组成,上面分布有叶绿素、藻胆色素等光合色素和有关酶类, 是光合作用的场所。③ 载色体 (chromatophore) 是一些不放氧的光合细菌的细胞质膜多次凹陷折叠而形成 的片层 状、微管状或囊状结构。载色体含有菌绿素和类胡萝卜素等光合色素及进行光合磷酸化所需要的酶类和电子传递体, 是进行光合作用的部位。④ 羧酶体 (carboxysome) 或称 多角体 (polyhedral body) 是自养细菌所特有的内膜结构。 羧酶体由以蛋白质为主的单层膜包围,厚约 35nm ,内含固定 CO 2 所需的 1,5- 二磷酸核酮糖羧化酶和 5- 磷酸核 酮糖激酶,是自养细菌固定 CO 2 的场所。 (三) 、细胞质 细胞质 (cytoplasm) 是细胞膜内的物质,除细胞核外皆为细胞质。它无色透明,呈粘胶状,主要成分为水、蛋白 质、核酸、脂类,也含有少量的糖和盐类。由于富含核酸,因而嗜碱性强,幼令菌着色均匀。此外,细胞质内还含有 核糖体、颗粒状内含物和气泡等物质。 1、核糖体 核糖体 (ribosome) 亦称核蛋白体,为多肽和蛋白质合成的场所。在电子显微镜下可见到细菌的核糖体游离于细 胞质中,系 70S 的颗粒,由 50S 和 30S 两个亚单元组成,化学成分为蛋白质与核糖核酸 (RNA) 。细菌细胞中绝大 部分 ( 约 90%) 的 RNA 存在于核糖体内。原核生物的核糖体常以游离状态或多聚核糖体状态分布于细胞质中。而真 核细胞的核糖体既可以游离状态存在于细胞质中,也可结合于内质网上。 2、内含物 很多细菌在营养物质丰富的时候,其细胞内聚合各种不同的贮藏颗粒,当营养缺乏时,它们又能被分解利用。这 种贮藏颗粒可在光学显微镜下观察到,通称为 内含物 ( cytoplasmic inclusions ) 。贮藏颗粒的多少可随菌令及培 养条件不同而改变。 异染颗粒 (metachromatic granules) 又称 捩转菌素 ( volutin ) ,最早发现于迂回螺 ( Spirillum volutans ) 中。异染粒是以无机偏磷酸盐聚合物为主要成分的一种无机磷的贮备物。异染颗粒嗜碱性或嗜中性较强,用蓝色染料 ( 如甲苯胺蓝或甲烯蓝 ) 染色后不呈蓝色而呈紫红色,故称异染颗粒。 聚 β - 羟基丁酸 ( poly- β -hydroxybutyric acid , PHB ) 颗粒 ,是一种碳源和能源性贮藏物。它是 D-3- 羟基丁酸的直链聚合物。用革兰氏染色时,这类物质不着色 ,但易被脂溶性染料如苏丹黑着色,在光学显微镜下可见 (图 1.11)。根瘤菌属(Rhizobium ) 、固氮菌属( Azotobacter ) 、假单胞菌属(Pseudomonas)等细菌常积累 PHB。 图 1.11 细胞内含物 ( Madigan et al., 2000) A. 细胞中的硫滴 B. 聚 β—羟基丁酸颗粒 肝糖粒 ( glycogen ) 和 淀粉粒 肝糖粒较小,只能 在电镜下观察到,如用稀碘液染色成红褐色,可在光学显 微镜下看到。有的细菌积累淀粉粒,用碘液可染成深蓝色。 肝糖粒、淀粉粒都是碳源贮藏物。 硫滴 ( sulfur globules ) 某些氧化硫的细菌细 胞内可积累硫滴。如贝氏硫菌属 ( Beggiatoa ) 、发硫 菌属 ( Thiothrix ) 在细胞内常含有强折光性的硫滴, 此为贮存的硫,系通过氧化硫化氢而形成,作为能量储备, 需要时可被细菌再利用。 磁小体 ( megnetosome ) 存在于水生细菌和趋磁细菌中,是细胞内磁铁矿 Fe 3 O 4 的晶体颗粒,数目不等。 不同种类的细菌磁小体形态不同,有正方形、长方形、还有刺状之分。含有磁小体的细菌表现出趋磁性、即沿着地磁 场转向和迁移。磁小体由一层含有磷脂、蛋白质和糖蛋白的膜包围。 3 、气泡 某些水生细菌,如蓝细菌、不放氧的光合细菌和盐细菌细胞内贮存气体的特殊结构称气泡。气泡由许多小的的气 囊 (gas vesicle) 组成,气囊膜只含蛋白质而无磷脂。气泡的大小、形状和数量随细菌种类而异。气泡能使细胞保持 浮力,从而有助于调节并使细菌生活在它们需要的最佳水层位置,以利获得氧、光和营养。 ( 四 ) 、细胞核 细菌细胞的核位于细胞质内,无核膜、无核仁,仅为一核区,因此称为原始形态的核 (primitive form nucleus) 或拟核 (nucleoid) 。细菌细胞的原核只有一个染色体,主要含有具有遗 传特征的脱氧核糖核酸 (DNA) 。拟核中尚

有皇结和蛋白质。但没有真核生物细跑核所含的组蛋白人结构蛋白)·染色体是由双螺旋的大分子链构成 物总长度为不因) 3mm 例 coli的 不则 分子 3X 10 9 Da ,约有5×10 则可见1 2个核区。 大多数的遗传 是细茵生长发 代谢和遗传变异的控制中 车细菌中,除染色体DNA外,还存在一种能自我复制的 DN分 质粒分子量较细 茵染色体小,约2100×106Da。每个菌体内可有一至数个质粒。 同质粒的基因之间可发生重组,质粒基 因与染色体基因也可重组。质粒对细菌的生存并不是必需的,它可在菌体内自行清失,也可经一定处理后从细南中除 去,但不影响细南的生存。 不同的所粒分别含有使细菌且有某些特殊性状的基因,如致有性、抗药性、产生抗生素 降解某些化学物质(见表1-4)等。 质粒可以独立于染色体而转移,通过接合、转化或转导等方式可从一个菌体转入另一茵体。因此在遗传工程中可 以将细茵质粒作为基因的运载工具,构建新菌株。 表1一4细菌质拉所能控制的性状 (五) 细菌质粒 性罚 大肠杆茵中的F因子 使宿主产生性纤毛,决定细菌的“性别” 据这层粘性物质的厚度 可溶性及在细 肠杆茵中的Co1因厅 产生 -种蛋白质类杀菌素 一大肠杆茵索 丧面存在的状况可把它们分为荚膜 微英膜或粘液层。如果这层物质粘滞性 大肠杆茵中的Ent因子 立生肠毒素 较大,相对稳定地附若在细胞壁外,具 大肠杆茵中的Hy因了 决定溶血素的产 一定外形,厚约200mm,称为英膜 大肠杆菌中的RYB质粒 可使宿主产生限制性内切酶与甲基化酶 (Capsule)或大荚膜 年多G-细南中的R因了 抗胺药与多种抗生素 它与细胞结合力转 金黄色萄萄球菌中的P1258因子抗Cd+ Hg+等重金属离子 细胞表面除去 金苗色萄萄球菌中的PA10 产生内毒素B 假单胞菌中的质粒 能降解某些复杂的有机物 图L.12荚膜Madi 限癌病农杆菌中的T1质粒 能使感垫梢株产生根席病 et 1 u1e)的厚度在200nm以下,它与细朐表面结 较紧,用光学显微镜不易观察到,但可采用血清学方法证明其存在。微 膜易被胰蛋白酶消化。 粘液层(slime layer)比荚膜疏松,无明显形状,悬浮在基质中更 易溶解,并能增加培养基粘度。 通常情况下,每个菌体外面包围一层荚膜。但有的细菌,它们的荚 膜物 起成为 物质的醇处理,也不会杀死细菌】 膜的主要成分因菌种而异, 大多为多糖 明,称光滑型S因 多肽或蛋白质,也含有一些其它成分 产荚膜的细菌茵落通常光滑透 荚膜的主要作用是作为细胞外碳源和能源性贮藏物质,并能保护细胞免受干燥的影响,同时能增强某些病原菌的 致病能力,使之抵抗宿主吞噬细胞的吞噬。例如能引起肺炎的肺炎双球菌型,如果失去了荚膜,则成为非致病菌。 有些产荚膜细菌,如肠膜明串珠菌(Leuconostoc mesenteroides)则可用于葡聚糖的工业生产,葡聚糖已被用 来治疗失血性休克的血浆代用品。野油莱黄单胞菌(anthomonas campestris)的粘液层的胞外多糖一黄原胶已被 用于石油开采中的钻井液添加剂以及印染和食品等工业中。而菌胶团则在污水生物处理中对活性污泥的形成、作用与 沉降性能等均具直要影。 牛奶、蜜糖、面包及其它含糖液变得“粘胶状”就是 由于 细茵的 细胞表面伸出细长、波曲、毛发状的附属物称为鞭毛( a 长度常为细胞的若干倍, 层长可70 因此,用光学显微镜看不见。 如果采用特殊的鞭毛染色法,使染料沉积在鞭毛上,加粗其直径,就可在光学显微镜下观察到细菌鞭毛 但真实形态只有在电镜下可见(见图1.13)。另外,采用悬滴法及暗视野映光法观察细菌运动状态 及用半固体琼脂穿刺培养,从细菌生长的扩散情况,可初步判断细菌是否有鞭毛。 细菌鞭毛的数目和者生位置是细菌种的特征。据此,可将有鞭毛的细菌分为四类(图1.14):① 端单毛菌(nonotrichaete)在菌体的一端只生一根鞭毛,如霍乱弧菌(Vibrio cholerae)。②两端单鞭毛菌 (amphitrichaete)菌体两端各具一根鞭毛,如鼠咬热螺旋体(Spirochaet a morsusmris)。③丛生鞭毛菌 1 ophotrichaete)菌体 如铜绿假单胞菌(Pse aeruginose 菌体两端各具一束鞭毛,如红 色螺菌(Soirillun tubr圆。O周生鞭毛随(peritrichact⊙周身都有题毛,知大肠杆商,枯草杆商穿
有少量 RNA 和蛋白质。但没有真核生物细胞核所含的组蛋白 ( 结构蛋白 ) 。染色体是由双螺旋的大分子链构成,一 般呈环形结构,总长度为 0. 25 ~ 3mm (例如 E.coli 的 DNA 长约 1mm ,分子量为 3 × 10 9 Da ,约有 5 × 10 6 个 bp, 至少含 5 × 103 个基因)。拟核在静止期常呈球形或不规则的棒状或哑铃形。一个细菌在正常情况下只有 一个核区,而细菌处于活跃生长时,由于 DNA 的复制先于细胞分裂,一个菌体内往往有 2~4 个核区,低速率生长时, 则可见 1 ~ 2 个核区。原核携带了细菌绝大多数的遗传信息,是细菌生长发育、新陈代谢和遗传变异的控制中心。 在细菌中,除染色体 DNA 外,还存在一种能自我复制的小环状 DNA 分子,称质粒 (plasmid) 。质粒分子量较细 菌染色体小,约 2 ~ 100 × 10 6 Da 。每个菌体内可有一至数个质粒。 不同质粒的基因之间可发生重组,质粒基 因与染色体基因也可重组。质粒对细菌的生存并不是必需的,它可在菌体内自行消失,也可经一定处理后从细菌中除 去,但不影响细菌的生存。不同的质粒分别含有使细菌具有某些特殊性状的基因,如致育性、抗药性、产生抗生素、 降解某些化学物质 ( 见表 1-4) 等。 质粒可以独立于染色体而转移,通过接合、转化或转导等方式可从一个菌体转入另一菌体。因此在遗传工程中可 以将细菌质粒作为基因的运载工具,构建新菌株。 表 1-4 细菌质粒所能控制的性状 (五) 、荚膜 有些细菌生活在一定营养条件下, 可向细胞壁外分泌出一层粘性物质,根 据这层粘性物质的厚度,可溶性及在细 胞表面存在的状况可把它们分为荚膜、 微荚膜或粘液层。如果这层物质粘滞性 较大,相对稳定地附着在细胞壁外,具 一定外形,厚约 200nm ,称为荚膜 (Capsule) 或大荚膜 (macrocapsule) 。 它与细胞结合力较 差。通过液体震荡培养或离心可将其从 细胞表面除去。荚膜很难着色,用负染 色法可在光学显微镜下观察到,即背景 和细胞着色,荚膜不着色 ( 图 1.12)。 图 1.12 荚膜(Madigan et al.,2000) 微荚膜 (microcapsule) 的厚度在 200nm 以下,它与细胞表面结合 较紧,用光学显微镜不易观察到,但可采用血清学方法证明其存在。微荚 膜易被胰蛋白酶消化。 粘液层 (slime layer) 比荚膜疏松,无明显形状,悬浮在基质中更 易溶解,并能增加培养基粘度。 通常情况下,每个菌体外面包围一层荚膜。但有的细菌,它们的荚 膜物质互相融合。在一起成为一团胶状物,称菌胶团( zoogloea ),其 内常包含有多个菌体。荚膜产生受遗传特性控制,但并非是细胞绝对必要 的结构,失去荚膜的变异株同样正常生长。而且,即使用特异性水解荚膜 物质的酶处理,也不会杀死细菌。 荚膜的主要成分因菌种而异,大多为多糖、多肽或蛋白质,也含有一些其它成分。 产荚膜的细菌菌落通常光滑透 明,称光滑型 (S 型 ) 菌落,不产荚膜细菌菌落表面粗糙,称粗糙型 (R 型 ) 菌落。 荚膜的主要作用是作为细胞外碳源和能源性贮藏物质,并能保护细胞免受干燥的影响,同时能增强某些病原菌的 致病能力,使之抵抗宿主吞噬细胞的吞噬。例如能引起肺炎的肺炎双球菌Ⅲ型,如果失去了荚膜,则成为非致病菌。 有些产荚膜细菌,如肠膜明串珠菌 ( Leuconostoc mesenteroides ) 则可用于葡聚糖的工业生产,葡聚糖已被用 来治疗失血性休克的血浆代用品。野油菜黄单胞菌( Xanthomonas campestris )的粘液层的胞外多糖—黄原胶已被 用于石油开采中的钻井液添加剂以及印染和食品等工业中。而菌胶团则在污水生物处理中对活性污泥的形成、作用与 沉降性能等均具重要影响。产荚膜细菌,常常给生产带来麻烦。牛奶、蜜糖、面包及其它含糖液变得“粘胶状”就是 由于受了某些产荚膜细菌的污染。有些细菌能籍荚膜牢固地粘附在牙齿表面引起龋齿。 ( 六 ) 、鞭毛 图 1.13 细菌鞭毛的电镜照片 某些细菌的细胞表面伸出细长、波曲、毛发状的附属物称为鞭毛 (flagella) 。 鞭毛细而长,其 长度常为细胞的若干倍,最长可达 70 μ m ,但直径只有 10 ~ 20nm 。因此,用光学显微镜看不见。 如果采用特殊的鞭毛染色法,使染料沉积在鞭毛上,加粗其直径,就可在光学显微镜下观察到细菌鞭毛, 但真实形态只有在电镜下可见 ( 见图 1.13 ) 。另外,采用悬滴法及暗视野映光法观察细菌运动状态 及用半固体琼脂穿刺培养,从细菌生长的扩散情况,可初步判断细菌是否有鞭毛。 细菌鞭毛的数目和着生位置是细菌种的特征。据此,可将有鞭毛的细菌分为四类(图 1.14):①一 端单毛菌(monotrichaete)在菌体的一端只生一根鞭毛,如霍乱弧菌(Vibrio cholerae)。②两端单鞭毛菌 (amphitrichaete)菌体两端各具一根鞭毛,如鼠咬热螺旋体(Spirochaet a morsusmuris)。③丛生鞭毛菌 (lophotrichaete)菌体一端生一束鞭毛,如铜绿假单胞菌(Pseudomonas aeruginosa);菌体两端各具一束鞭毛,如红 色螺菌(Spirillum rubrum)。④周生鞭毛菌(peritrichaete)周身都有鞭毛,如大肠杆菌、枯草杆菌等。 细菌质粒 性状 大肠杆菌中的 F 因子 使宿主产生性纤毛,决定细菌的“性别” 大肠杆菌中的 Col 因子 产生一种蛋白质类杀菌素—大肠杆菌素 大肠杆菌中的 Ent 因子 产生肠毒素 大肠杆菌中的 Hty 因子 决定溶血素的产生 大肠杆菌中的 RYB 质粒 可使宿主产生限制性内切酶与甲基化酶 许多 G - 细菌中的 R 因子 抗磺胺药与多种抗生素 金黄色葡萄球菌中的 P 1258 因子 抗 Cd ++ 、 Hg ++ 等重金属离子 金黄色葡萄球菌中的 PZA10 产生内毒素 B 假单胞菌中的质粒 能降解某些复杂的有机物 根癌病农杆菌中的 Ti 质粒 能使感染植株产生根癌病
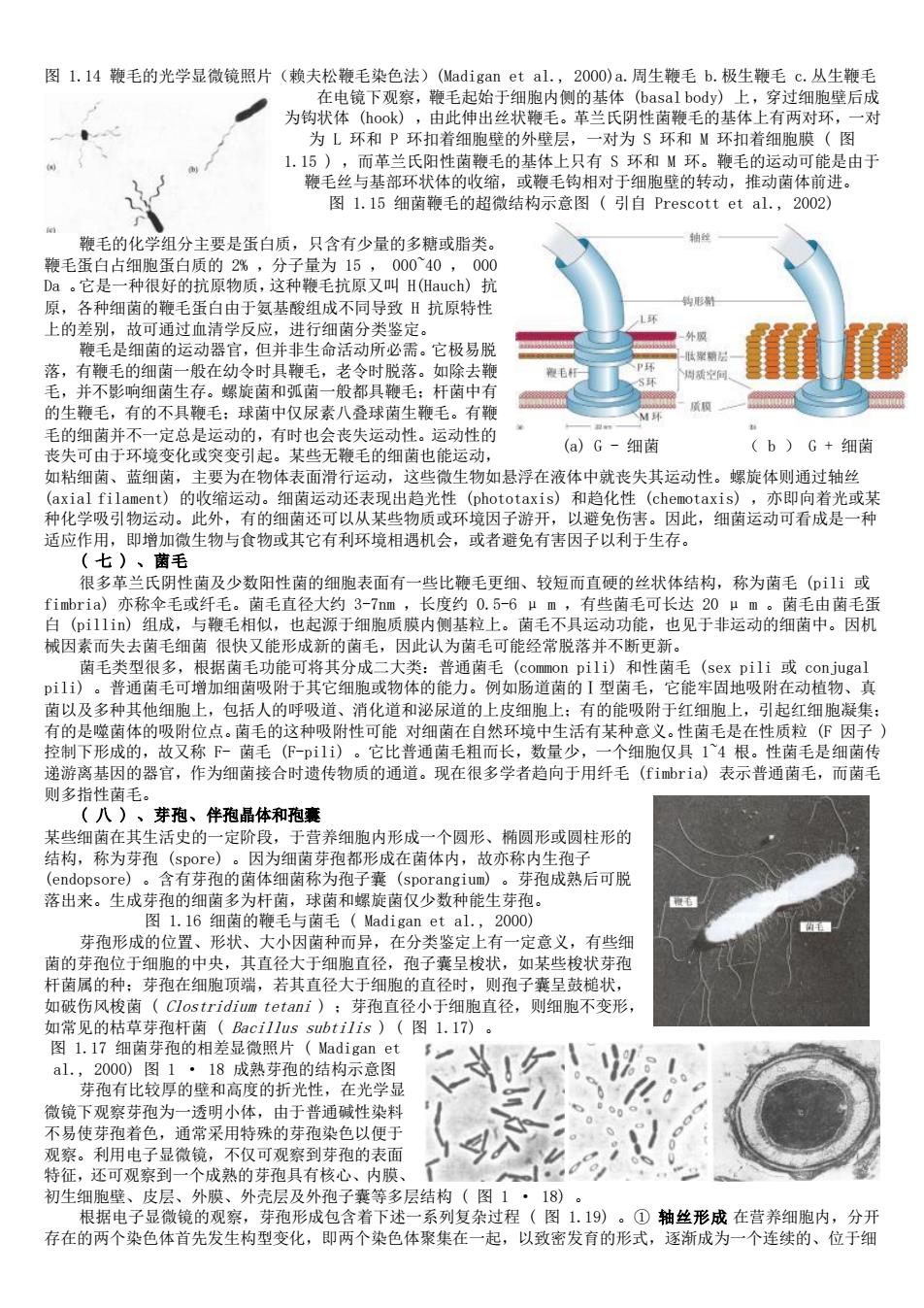
图L.14鞭毛的光学显微镜照片(赖夫松鞭毛染色法)(Madigan et al.,2000)a.周生鞭毛b,极生鞭毛C,从生鞭毛 为钩状 镜下观察:毛起始织空侧的基使阴性萄神的 胞壁后成 环和P环 115 的 不和M 毛丝与基部环状休的 的使毛有对于壁的转 能是由于 体 图1.15细菌鞭毛的超微结构示意图(引自Prescott et al.,2002) 鞭毛的化学组分主要是蛋白质, 只含有少量的名糖或脂类 鞭毛蛋白占细胞蛋白质的2% 分于量为1 00040, 00 种很好的抗原物局 抗原又叫H(Hauch) 星 各种细菌的鞭毛 司导致日抗原特性 如 都且毛:杆南中 的生鞭毛,有的不具鞭毛:球菌中仅尿素八叠球菌生鞭毛。有粮 手的细菌并不一定总是运动的,有时也会丧失运动性。运动性的 丧失可由于环境变化或突变引起。某些无鞭毛的细菌也能运动, (a)G-细菌 (b)G+细菌 如粘细菌、蓝细菌,主要为在物体表面滑行运动,这些微生物如悬浮在液体中就丧失其运动性。螺旋体则通过轴丝 学吸引物的收缩运动。细运动还表现出自光性oo1和自化性(cherotaxi (axial fila 亦即间若光或果 此外 图还可以从果些彩 或环境因子游开,以避免伤害。因此,细茵运动可看成是一种 适应 生物与食物或其它有利环境相遇机会,或者避免有害因子以利于生存。 七) 毛 很多氏 性菌及少数阳性菌的细胞表面有 a)亦称伞毛或红 称为商毛61 白(im)组成,与鞭毛相似,也起源于细胞质膜内侧基粒上。菌毛不具运动功能, 械因素而失去菌毛细菌很快又能形成新的菌毛,因此认为菌毛可能经常脱落并不断更新。 菌毛类型很多,根据菌毛功能可将其分成二大类:普通菌毛(common pili)和性菌毛(sex pili或conjugal p1)。普通菌毛可增加细菌吸附于其它细胞或物体的能力。例如肠道菌的I型菌毛,它能牢固地吸附在动植物、真 茵以及多种其他细胞上,包括人的呼吸道、消化道和泌尿道的上皮细胞上:有的能吸附于红细胞上,引起红细胞凝集 有的是噬菌体的吸附位点。菌毛的这种吸附性可能对细菌在自然环境中生活有某种意义。性菌毛是在性质粒(『因子 (F-p1l1) 细胞仅具1`4根。性菌毛是细菌 作为细菌接合时遗传物质的通道。现在很多学者趋向于用纤毛山)表示普通菌毛,而商 (八)、芽孢、伴孢品体和孢囊 定阶段 王细购内形成 一个圆形、椭圆形或圆柱形的 称为葬孢 )。因为细菌芽孢都形成在茵体内 故亦称内生孢 含有芽孢的茵体细菌称为孢子囊(sporangium 芽孢成熟后可脱 落出来。生成芽孢的细菌多为杆菌,球菌和螺旋菌仅少数种能生芽 图1.16细菌的鞭毛与菌毛(Madigan et al.,200o) 芽孢形成的位置、形状、大小因菌种而异,在分类鉴定上有一定意义,有些细 茵的芽孢位于细胞的中央,其直径大于细胞直径,孢子囊呈梭状,如某些梭状芽孢 轩菌属的种: 芽孢在细胞项瑞, 若其直径大于细胞的直径时, 则孢子囊呈鼓桅状 芽孢直 小于细胞直径,则细胞不变形 1.7 2000)图1·18成熟芽孢的结构示意图 芽孢有比较厚的壁和高度的折光性,在光学显 微镜下观察芽孢为一透明小体,由于普通碱性染料 不易使芽孢着色,通常采用特殊的芽孢染色以便于 观察。利用电子显微镜,不仅可观察到芽孢的表面 <入 特征,还可观察到 个成熟的芽孢具有核心、内膜 初生细胞壁 外膜、外壳层及外孢子囊等多层结构(图 18) 杂过程 19 集 一起
图 1.14 鞭毛的光学显微镜照片(赖夫松鞭毛染色法)(Madigan et al., 2000)a.周生鞭毛 b.极生鞭毛 c.丛生鞭毛 在电镜下观察,鞭毛起始于细胞内侧的基体 (basal body) 上,穿过细胞壁后成 为钩状体 (hook) ,由此伸出丝状鞭毛。革兰氏阴性菌鞭毛的基体上有两对环,一对 为 L 环和 P 环扣着细胞壁的外壁层,一对为 S 环和 M 环扣着细胞膜 ( 图 1.15 ) ,而革兰氏阳性菌鞭毛的基体上只有 S 环和 M 环。鞭毛的运动可能是由于 鞭毛丝与基部环状体的收缩,或鞭毛钩相对于细胞壁的转动,推动菌体前进。 图 1.15 细菌鞭毛的超微结构示意图 ( 引自 Prescott et al., 2002) 鞭毛的化学组分主要是蛋白质,只含有少量的多糖或脂类。 鞭毛蛋白占细胞蛋白质的 2% ,分子量为 15 , 000~40 , 000 Da 。它是一种很好的抗原物质,这种鞭毛抗原又叫 H(Hauch) 抗 原,各种细菌的鞭毛蛋白由于氨基酸组成不同导致 H 抗原特性 上的差别,故可通过血清学反应,进行细菌分类鉴定。 鞭毛是细菌的运动器官,但并非生命活动所必需。它极易脱 落,有鞭毛的细菌一般在幼令时具鞭毛,老令时脱落。如除去鞭 毛,并不影响细菌生存。螺旋菌和弧菌一般都具鞭毛;杆菌中有 的生鞭毛,有的不具鞭毛;球菌中仅尿素八叠球菌生鞭毛。有鞭 毛的细菌并不一定总是运动的,有时也会丧失运动性。运动性的 丧失可由于环境变化或突变引起。某些无鞭毛的细菌也能运动, 如粘细菌、蓝细菌,主要为在物体表面滑行运动,这些微生物如悬浮在液体中就丧失其运动性。螺旋体则通过轴丝 (axial filament) 的收缩运动。细菌运动还表现出趋光性 (phototaxis) 和趋化性 (chemotaxis) ,亦即向着光或某 种化学吸引物运动。此外,有的细菌还可以从某些物质或环境因子游开,以避免伤害。因此,细菌运动可看成是一种 适应作用,即增加微生物与食物或其它有利环境相遇机会,或者避免有害因子以利于生存。 ( 七 ) 、菌毛 很多革兰氏阴性菌及少数阳性菌的细胞表面有一些比鞭毛更细、较短而直硬的丝状体结构,称为菌毛 (pili 或 fimbria) 亦称伞毛或纤毛。菌毛直径大约 3-7nm ,长度约 0.5-6 μ m ,有些菌毛可长达 20 μ m 。菌毛由菌毛蛋 白 (pillin) 组成,与鞭毛相似,也起源于细胞质膜内侧基粒上。菌毛不具运动功能,也见于非运动的细菌中。因机 械因素而失去菌毛细菌 很快又能形成新的菌毛,因此认为菌毛可能经常脱落并不断更新。 菌毛类型很多,根据菌毛功能可将其分成二大类:普通菌毛 (common pili) 和性菌毛 (sex pili 或 conjugal pili) 。普通菌毛可增加细菌吸附于其它细胞或物体的能力。例如肠道菌的Ⅰ型菌毛,它能牢固地吸附在动植物、真 菌以及多种其他细胞上,包括人的呼吸道、消化道和泌尿道的上皮细胞上;有的能吸附于红细胞上,引起红细胞凝集; 有的是噬菌体的吸附位点。菌毛的这种吸附性可能 对细菌在自然环境中生活有某种意义。性菌毛是在性质粒 (F 因子 ) 控制下形成的,故又称 F- 菌毛 (F-pili) 。它比普通菌毛粗而长,数量少,一个细胞仅具 1~4 根。性菌毛是细菌传 递游离基因的器官,作为细菌接合时遗传物质的通道。现在很多学者趋向于用纤毛 (fimbria) 表示普通菌毛,而菌毛 则多指性菌毛。 ( 八 ) 、芽孢、伴孢晶体和孢囊 某些细菌在其生活史的一定阶段,于营养细胞内形成一个圆形、椭圆形或圆柱形的 结构,称为芽孢 (spore) 。因为细菌芽孢都形成在菌体内,故亦称内生孢子 (endopsore) 。含有芽孢的菌体细菌称为孢子囊 (sporangium) 。芽孢成熟后可脱 落出来。生成芽孢的细菌多为杆菌,球菌和螺旋菌仅少数种能生芽孢。 图 1.16 细菌的鞭毛与菌毛 ( Madigan et al., 2000) 芽孢形成的位置、形状、大小因菌种而异,在分类鉴定上有一定意义,有些细 菌的芽孢位于细胞的中央,其直径大于细胞直径,孢子囊呈梭状,如某些梭状芽孢 杆菌属的种;芽孢在细胞顶端,若其直径大于细胞的直径时,则孢子囊呈鼓槌状, 如破伤风梭菌 ( Clostridium tetani ) ;芽孢直径小于细胞直径,则细胞不变形, 如常见的枯草芽孢杆菌 ( Bacillus subtilis ) ( 图 1.17) 。 图 1.17 细菌芽孢的相差显微照片 ( Madigan et al., 2000) 图 1 · 18 成熟芽孢的结构示意图 芽孢有比较厚的壁和高度的折光性,在光学显 微镜下观察芽孢为一透明小体,由于普通碱性染料 不易使芽孢着色,通常采用特殊的芽孢染色以便于 观察。利用电子显微镜,不仅可观察到芽孢的表面 特征,还可观察到一个成熟的芽孢具有核心、内膜、 初生细胞壁、皮层、外膜、外壳层及外孢子囊等多层结构 ( 图 1 · 18) 。 根据电子显微镜的观察,芽孢形成包含着下述一系列复杂过程 ( 图 1.19) 。① 轴丝形成 在营养细胞内,分开 存在的两个染色体首先发生构型变化,即两个染色体聚集在一起,以致密发育的形式,逐渐成为一个连续的、位于细 (a) G - 细菌 ( b ) G + 细菌

胞中央的轴丝状结构,并通过中间体与细胞膜相连接。有人认为这是芽孢早期所具有的特异结构,是抗辐射的物质基 隔膜形成在接近细 细胞膜内陷, 心延伸, 此同时的 两层极性 的细胞膜组成, 其中内膜将发有成为营养细胞的细胞膜, 皮层形成由于前孢子迅速进行合成作用 新形成的物质 F前孢子的两层极性相反的细胞膜之间 逐渐发有形成皮层( a2+的吸收,出现pe 与此同时,随若2,6-吡啶二羧酸 id,简称DPA)的合成, 复合物:而且在前孢子的外面也开始形成外 壳层。此时的皮层,似乎变成了一种呈现出条纹的多层结构。⑤孢子外光层的形成在皮层形成过程中,前孢子外膜 表面合成外壳物质,并沉积于皮层外表,逐渐形成一个连续的致密层。在外壳中含有非常多的半胱氨酸和疏水性氨基 受季是的数子凳胶袋《福品蔬务实全事充政。手具有了银强航热在和防味结向 图1.19细菌芽孢形成的几个阶段(Madigan et al.,1997) 成过程, 仅能见到以下变化: 的区 华性逐增强形成成熟 小时成 袋 芽孢没有繁殖意义 为 孢,而一个芽孢也只产生一个营养细胞。芽孢仅仅是芽孢细菌 生活史中的一环,是细菌的休眠体。 形成芽孢需要一定的外界条件,这些条件因菌种而异。然 而,芽孢一旦形成,则对恶劣环境条件均具有很强的抵抗能力。 有的芽孢,在定条件下可保存十年而不丧失其生活力芽 孢尤其能耐高温 草杆菌的芽孢在 小时, 破伤风 的芽孢口 极菌的芽恒则 铜射 天多数化学杀菌剂也具 有极大的抗性 构和化学特性有关(见表1-5) 表1-5细菌的芽孢和营养细胞的区别 项目 营养胞孢 典型革兰氏阳府髓、孢子衣、外壁 光学显微镜下 无折光性 有折光性 化学组成: 什空一街酸 聚B一羟基丁酸(PHB) 名边 蛋白质 较高 含硫氨基酸 酶活性 代谢作用(02利用) 低或无 大分子合成 RNA 低或无 抗热性 抗幅射 抗药和酸 染色力 需用特殊方法染色
胞中央的轴丝状结构,并通过中间体与细胞膜相连接。有人认为这是芽孢早期所具有的特异结构,是抗辐射的物质基 础。② 隔膜形成 在接近细胞一端处,细胞膜内陷,向心延伸,产生隔膜,将细胞分成大小两部分。与此同时,轴丝 状结构也分为两部分。③ 前孢子形成 在细胞中,较大部分的细胞膜围绕较小部分迅速地继续延伸,直至将小的部分 完全包围到大的部分中为止 ? 这个新形成的细胞 ( 结构 ) 称为前孢子 (forespore) 。此时前孢子由两层极性相反 的细胞膜组成,其中内膜将发育成为营养细胞的细胞膜。④ 皮层形成 由于前孢子迅速进行合成作用,新形成的物质 沉积于前孢子的两层极性相反的细胞膜之间,逐渐发育形成皮层 (corter) 。与此同时,随着 2,6- 吡啶二羧酸 (dipicolinic acid ,简称 DPA) 的合成, Ca 2+ 的吸收,出现 DPA-Ca 复合物;而且在前孢子的外面也开始形成外 壳层。此时的皮层,似乎变成了一种呈现出条纹的多层结构。⑤ 孢子外壳层的形成 在皮层形成过程中,前孢子外膜 表面合成外壳物质,并沉积于皮层外表,逐渐形成一个连续的致密层。在外壳中含有非常多的半胱氨酸和疏水性氨基 酸,并且继续积累 DPA 和 Ca 2+ 。⑥ 芽孢成熟 芽孢合成过程全部完成,此时芽孢具有了很强的抗热性和特殊结构。 ⑦ 芽孢的释放 孢子囊壁破裂 ( 溶解 ) ,释放出成熟的芽孢。 图 1.19 细菌芽孢形成的几个阶段( Madigan et al., 1997) 在光学显微镜下观察芽孢形成过程,仅能见到以下变化: 首先,在细胞一端出现一个折光性较强的区域,即前孢子阶段; 然后,折光性逐渐增强,形成成熟的孢子;几小时后,成熟的 芽孢部分地或全部脱离子囊壁而释放,呈游离状。 芽孢没有繁殖意义,因为一个细胞内一般只形成一个芽 孢,而一个芽孢也只产生一个营养细胞。芽孢仅仅是芽孢细菌 生活史中的一环,是细菌的休眠体。 形成芽孢需要一定的外界条件,这些条件因菌种而异。然 而,芽孢一旦形成,则对恶劣环境条件均具有很强的抵抗能力。 有的芽孢,在一定条件下可保存几十年而不丧失其生活力。芽 孢尤其能耐高温,如枯草杆菌的芽孢在沸水中可存活 1 小时, 破伤风杆菌的芽孢可存活 3 小时,而肉毒梭菌的芽孢则可忍 受 6 小时左右,即使在 180 ℃的干热中仍可存活 10 分钟。 除耐热外,芽孢也能抵抗低温,它在液氮温度 (-190 ℃ ) 中 6 个月仍能存活。芽孢对辐射、干燥和大多数化学杀菌剂也具 有极大的抗性。芽孢之所以具有如此高度的抗逆性,这与其结 构和化学特性有关 ( 见表 1-5) 。 表 1-5 细菌的芽孢和营养细胞的区别 项目 营养细胞 芽孢 结构 典型革兰氏阳性 有髓、孢子衣、 外壁 光学显微镜下 无折光性 有折光性 化学组成: 钙 低 高 吡啶二羧酸 无 有 聚 β—羟基丁酸(PHB) 有 无 多糖 高 低 蛋白质 较低 较高 含硫氨基酸 低 高 酶活性 高 低 代谢作用 (O 2 利用 ) 高 低或无 大分子合成 有 无 mRNA 有 低或无 抗热性 低 高 抗幅射 低 高 抗药和酸 低 高 染色力 可染 需用特殊方法染色

①芽孢的一个显著特点是游离水含量远低于营养 溶菌酶作用 ]感 佰抗性 细胞,使核酸和蛋白质不易变性。芽孢的酶组成型与营养 是附着在颗 ②芽孢另 存 二羧酸在芽孢中以钙盐形式 占细菌芽孢于重的5 院一象的 程中,随若DPA的形成而具抗热性,芽孢萌发时毗啶二羧酸又释放至培养基中,同时也丧失其抗热性。显然DPA与 芽孢的抗热性有关。催化DPA合成中最后阶段的酶也是芽孢形成所特有的,由于DPA钙盐的存在可改变酶的构型 使酶对热相当稳定。 ③芽孢在适合的条件下可萌发。适合芽孢萌发的条件包括水和营养物质,适合的温度,氧浓度以及某些必须的条 件。加热到80℃`85℃处理几分钟可促进芽孢萌发,芽孢萌发时首先丧失抗性、折光性和增加可染性。芽孢萌发时 开始吸收水分、盐类和营养物质而体积膨大, 与此同时,耐热力和折光性逐渐降低:在细胞质内部发生了 切提高 呼吸 白质 皮等的合成,因 使细 thu 在形成的同 90 )。它主要对鳞翅目 的昆虫有毒性,由于这种品体毒素对人畜毒性很低,故国内外均以工业化方式大量生产菌剂,以杀死某些农业害虫。 图1.20苏云金杆菌的芽孢和件孢晶体 孢囊细菌除产生芽孢作为休眠体以抵抗不良生活环境外,某些细前 还能形成其他休眠构造:如固氨菌的孢囊(cyst),粘球菌的粘液孢子 。固氯茵的孢囊(图 为球型或卵圆形,中心为一稠密的中心体 (central body) 8 羟基丁酸颗粒PB· 度小 大月 维氏固氮菌 忌微 照相表明孢子外壁是由 △而 70 孢子外壁的化学组分为32碳水化合物, 28%蛋白质 30%脂类和32% 分。孢子内壁的化学组分为44%碳水化合物,9.1%蛋白质,37%脂类和4.1%灰分。 图1.21固氮菌的营养细胞、孢囊、孢囊结构 A.营养细胞B.孢囊(Madigan et al.,2000)C.圆褐固氮茵孢囊的超薄切片电镜照片(浙江大学学报,2001) 固氨菌的孢囊具有抗干燥、抗机械破坏 0 抗电离辐射的作用,但并不特别抗热,也不完 外源性的能源。孢囊的形 分学交化是运动的杆状细 转变成不运动球状细胞,数小时后壁增厚,并 逐步发育成有折光性的孢囊。随若孢囊的成 C 熟,固氮菌丧失其固氮能力。在适宜条件下孢 囊可萌发。萌发时中心体膨大,孢囊内壁消失,孢囊外壁出现断裂,最后从崩溃的孢囊结构中,生出年幼的营养细胞。 三、细菌的繁殖及其群体特征 )、细菌的繁殖 个子细胞 称作司 现为细胞的横分 ,称为裂殖 绝大多数类群在分裂时产 大的和形相的 过程 首先是核的分裂和隔膜的形成。细菌染色体的复制往往先于细胞分裂,并随者细菌生长而分开。与此同时 细胞赤道附近的细胞膜从外向中心作环状推进,然后闭合形成 个垂直于细胞长轴的细胞质路膜, 铺细跑质知细胞起 的一分为 。第二步形成横隔壁,如状芽孢杆菌(阳cillus m oides),随若细胞膜的向内陷,母细胞的细胞壁 也跟着由四周向中心逐渐延伸,把细胞质隔膜分为两层,每层分别成为子细胞的细胞膜,横隔壁也逐渐分为两层,这 样每个细胞便各自具备了 个完整的细胞壁。有的细菌如链球菌、双球菌等在分裂过程中,横隔壁尚未完全形成,细 胞就停止了生长,留下了一个小孔,此时两个细胞的细胞膜仍然相连,即形成了“胞间连丝” 第二步是子细胞分离 有些种类的细菌细胞,在横隔壁形成后不久便相互分开,呈单个游离状态:而有的却数个细胞相连呈短链状或多个邦 列成长链状。尤其是球菌,因分裂面的不同,使分裂后排列成单球菌、双球菌、链球菌、四联球菌、八叠球菌和葡萄
① 芽孢的一个显著特点是游离水含量远低于营养 细胞,使核酸和蛋白质不易变性。芽孢的酶组成型与营养 细胞也有差别,芽孢只含有少量酶,并处于不活跃状态。芽孢的抗热性也与芽孢内具抗热性的酶有关。例如,营养细 胞中的过氧化氢酶是可溶的和热敏感性的,而芽孢中的过氧化氢酶则是附着在颗粒上和热抗性的。而且两种酶的动力 学和血清学特性均不同。 ② 芽孢另一独特之处是含有 2, 6- 吡啶二羧酸 (dipicolinic acid , DPA) 。吡啶二羧酸在芽孢中以钙盐形式 存在,占细菌芽孢干重的 5 %~ 15% 。在细菌营养细胞及其他生物细胞中均未发现有吡啶二羧酸的存在。芽孢形成 过程中,随着 DPA 的形成而具抗热性,芽孢萌发时吡啶二羧酸又释放至培养基中,同时也丧失其抗热性。显然 DPA 与 芽孢的抗热性有关。催化 DPA 合成中最后阶段的酶也是芽孢形成所特有的,由于 DPA 钙盐的存在可改变酶的构型, 使酶对热相当稳定。 ③ 芽孢在适合的条件下可萌发。适合芽孢萌发的条件包括水和营养物质,适合的温度,氧浓度以及某些必须的条 件。加热到 80 ℃ ~85 ℃处理几分钟可促进芽孢萌发,芽孢萌发时首先丧失抗性、折光性和增加可染性。芽孢萌发时 开始吸收水分、盐类和营养物质而体积膨大,与此同时,耐热力和折光性逐渐降低;在细胞质内部发生了一系列生理 变化,着色力增强 ,酶活力提高,呼吸作用加强,可看到核分裂, DPA 和钙复合物外流,对外界各种不良因素的抵 抗力降低;随之肽聚糖分解,孢子囊壁破裂,皮层迅速破坏,长出芽管,逐渐发育成新的营养细胞。芽孢萌发过程中 还伴随有大分子细胞物质如 RNA 、蛋白质、 DNA 等的合成,因此,使细胞不断长大。 伴孢晶体 芽孢杆菌属有些种如苏云金芽孢杆菌 ( Bacillus thuringiensis ) ,在形成芽孢的同时,在细胞内产 生一颗菱形或双椎形的碱性蛋白晶体,称为伴孢晶体 (spore companioned crystal)( 图 1. 20) 。它主要对鳞翅目 的昆虫有毒性,由于这种晶体毒素对人畜毒性很低,故国内外均以工业化方式大量生产菌剂,以杀死某些农业害虫。 图 1.20 苏云金杆菌的芽孢和伴孢晶体 孢囊 细菌除产生芽孢作为休眠体以抵抗不良生活环境外,某些细菌 还能形成其他休眠构造:如固氮菌的孢囊 (cyst) ,粘球菌的粘液孢子 (myxospore) ,蛭弧菌的蛭孢囊 (bdellocyst) 等。固氮菌的孢囊 ( 图 1.20) 为球型或卵圆形,中心为一稠密的中心体 (central body) ,中 心体中往往含有数个折射颗粒,为聚 β - 羟基丁酸颗粒 (PHB) 。围绕 中心体的为两层厚薄不一的壁,内层称孢子内壁 (intine ,简写 in) , 密度小,宽而均匀。外层称孢子外壁 (exine 简写 ex) ,密度大且坚硬, 为紧密多层膜片状结构。维氏固氮菌 ( Azotobacter vinelandii ) 孢囊超薄切片的电子显微照相表明孢子外壁是由 三层膜状物迭合而成,厚约 70~75nm 。孢子外壁的化学组分为 32% 碳水化合物, 28% 蛋白质、 30% 脂类和 3.2% 灰 分。孢子内壁的化学组分为 44% 碳水化合物,9.1% 蛋白质, 37% 脂类和 4.1% 灰分。 图 1.21 固氮菌的营养细胞、孢囊、孢囊结构 A. 营养细胞 B. 孢囊(Madigan et al., 2000) C. 圆褐固氮菌孢囊的超薄切片电镜照片(浙江大学学报 , 2001) 固氮菌的孢囊具有抗干燥、抗机械破坏、 抗电离辐射的作用,但并不特别抗热,也不完 全休眠,能迅速氧化外源性的能源。孢囊的形 成受某些化合物的诱导,如正丁醇、β - 羟 基丁酸盐和巴豆酸等能促使孢囊形成。孢囊的 形成与芽孢不同,它是由整个营养细胞转变而 来,而不是由部分细胞物质转变而成。孢囊形 成过程,最先的形态学变化是运动的杆状细胞 转变成不运动球状细胞,数小时后壁增厚,并 逐步发育成有折光性的孢囊。随着孢囊的成 熟,固氮菌丧失其固氮能力。在适宜条件下孢 囊可萌发。萌发时中心体膨大,孢囊内壁消失,孢囊外壁出现断裂,最后从崩溃的孢囊结构中,生出年幼的营养细胞。 三、细菌的繁殖及其群体特征 ( 一 ) 、细菌的繁殖 细菌一般进行无性繁殖,表现为细胞的横分裂,称为裂殖。绝大多数类群在分裂时产生大小相等和形态相似的两 个子细胞,称作同形裂殖。电镜研究表明,细菌分裂大致经过细胞核和细胞质的分裂、横隔壁的形成、子细胞分离等 过程。 首先是核的分裂和隔膜的形成。细菌染色体 DNA 的复制往往先于细胞分裂,并随着细菌生长而分开。与此同时, 细胞赤道附近的细胞膜从外向中心作环状推进,然后闭合形成一个垂直于细胞长轴的细胞质隔膜,使细胞质和细胞核 均一分为二。第二步形成横隔壁,如蕈状芽孢杆菌 ( Bacillus mycoides ) ,随着细胞膜的向内陷,母细胞的细胞壁 也跟着由四周向中心逐渐延伸,把细胞质隔膜分为两层,每层分别成为子细胞的细胞膜,横隔壁也逐渐分为两层,这 样每个细胞便各自具备了一个完整的细胞壁。有的细菌如链球菌、双球菌等在分裂过程中,横隔壁尚未完全形成,细 胞就停止了生长,留下了一个小孔,此时两个细胞的细胞膜仍然相连,即形成了“胞间连丝”。第三步是子细胞分离。 有些种类的细菌细胞,在横隔壁形成后不久便相互分开,呈单个游离状态;而有的却数个细胞相连呈短链状或多个排 列成长链状。尤其是球菌,因分裂面的不同,使分裂后排列成单球菌、双球菌、链球菌、四联球菌、八叠球菌和葡萄 溶菌酶作用 敏感 有抗性 A B C