
第一章原核微生物 一内容类要细西、故线商、黄细萌等技微牛物的形本、大小细胞的结执、成分与功能以及它们的做方和商移 特征 按生物的系统发有和16sRNA分析,细胞生物可分为细菌、古菌和真核生物,细菌和古菌同属于原核微生物, 细南有基本形态和特殊形态,细菌细胞的大小以μm度量。G一菌与G+菌的细胞壁在结构和成分上的差异 决定了革兰氏染色的结果:细菌中还存在着缺壁菌。细菌细胞膜是细胞代谢活动的中心,此外有些细菌还存在着细胞 内浪系统。细胞质中核蛋拍体是多和蛋白质合成的场所,果些细菌细胞质内有各司括美魔能内含窗王、年包牛的 物质为 递遗传信 丝状的G+原核微生物 形与能分为 落特征 养特生菌丝与孢子丝,可形成分生孢 16 S rRNA中核苷酸顺序等都与细菌中的不同,也与真核生物不同。目前古菌分为4个 类群,生长在独特的生态环境。 蓝细菌细胞内含有独特的内膜结构(内囊体)和特有的色素蛋白(藻胆蛋白),是能进行光合作用的原核微生物。枝 原体为无细跑壁的最小的原核微生物。立克次氏体和农原体都是专性细胞内寄生物,但它们的形态、大小、寄主各不 相间。 近代生物学把生物区分为细胞生物和非细胞生物两大类。细胞生物包括一切具有细胞形态的生物,按系统发有和16S )、古菌(Archaea,曾用Archaebacteria)和 ligan et al.,2000) 原核生物与真核生物的区别 原核生物包括古菌和细菌,与真核生物的区 别综合列于表1一 主要差异有(1)、原核生 物的遗传物质主要是以双螺旋DNA构成的一条染 细菌 古生菌 真核生物 色体(chromosome),仅形成一个核区,没有核膜 包围,无核仁,称为原核(nucleoid)或拟核,无 组蛋白与之相结合。真核生物的遗传物质以双螺前 DNA构成一条或一条以上的多条染色体君 象古生界 形成 一核膜包用, 生格球两 鞭毛 并有 有自己的DN 可白主复制 2) 原楼 细的细胞质由细胞时(。e11 句用 双金 并有细胞膜大量褶皱内陷入细胞质中形成中间体 或称为间体(mesosome))。不含其他分化明显的细 胞器(organelles)。真核生物细胞同样由细胞膜 包围,但不内陷,内含多种细胞器,如主要进行呼吸能量代谢的线粒体(mitochondria)和光合作用的叶绿体 (chloroplast) 各种细胞器有各自的膜包围,细胞器膜与细胞膜之间无直接关系。(3)、原核生物和真核生物 细跑的蛋 质合成 在核蛋白体 不同,原核生物的核蛋白体为705,而真核生物的核蛋白体为80S 古和细菌在细胞形态结物。生长蝼 、遗传物质存在方式等方面相类似,因而同属原 核生物。但在分子生物学水平上,古菌和细菌之间有明显差别,主要表现见表1一 从这些差异可见,古菌确是不仅 在细胞化学组成上更是在分子生物学水平上不同于同属于原核生物的细菌和真核生物的另一类特殊生物类群。 古菌(archaea))、细菌(bacteria)和真核生物(eucaryoutes)三域(urkingdoms)的概念是沃斯(Woese)及 其同事1977年根据对代表性细菌类群的16 S FRNA碱基序列进行广泛比较后提出的,认为生物界的发育并不是一个 由简单的原核生物发育到较完全、较复杂的真核生物的过程,而是明显存在着三个发育不同的基因系统,即古菌、细 茵和真核生物。并认为这三个基因系统几乎是同时从某一起点各自发有而来,这一起点即是至今仍不明确的一个原始 域观念已被广泛接 菌和古菌 同的生物 类群,但它们的细胞形态和结构却基本 物的形结先 是指一大类细胞不具核膜,也无核仁,只有核区,称为原核的单细胞生物。本章介绍原
第一章 原核微生物 内容提要: 本章介绍了细菌、放线菌、蓝细菌等原核微生物的形态、大小;细胞的结构、成分与功能以及它们的繁殖方式和菌落 特征。 按生物的系统发育和 16s rRNA 分析,细胞生物可分为细菌、古菌和真核生物,细菌和古菌同属于原核微生物。 细菌有基本形态和特殊形态,细菌细胞的大小以 μ m 度量。 G - 菌与 G + 菌的细胞壁在结构和成分上的差异 决定了革兰氏染色的结果;细菌中还存在着缺壁菌。细菌细胞膜是细胞代谢活动的中心,此外有些细菌还存在着细胞 内膜系统。细胞质中核蛋白体是多肽和蛋白质合成的场所,某些细菌细胞质内含有各具不同功能的内含物。原核中的 遗传物质为 DNA ,质粒也具有储存和传递遗传信息的功能。一些细菌有特殊结构,包括荚膜、鞭毛、菌毛、芽孢、伴 孢晶体、孢囊。细菌以裂殖方式繁殖,不同的细菌具有不同的菌落特征和液体培养特征。 放线菌是分枝丝状的 G + 原核微生物,根据菌丝形态与功能可分为基内菌丝、气生菌丝与孢子丝,可形成分生孢 子。放线菌有其独特的菌落特征。 古菌的细胞壁、细胞膜、 16S rRNA 中核苷酸顺序等都与细菌中的不同,也与真核生物不同。目前古菌分为 4 个 类群,生长在独特的生态环境。 蓝细菌细胞内含有独特的内膜结构(内囊体)和特有的色素蛋白(藻胆蛋白),是能进行光合作用的原核微生物。枝 原体为无细胞壁的最小的原核微生物。立克次氏体和衣原体都是专性细胞内寄生物,但它们的形态、大小、寄主各不 相同。 近代生物学把生物区分为细胞生物和非细胞生物两大类。细胞生物包括一切具有细胞形态的生物,按系统发育和 16S rRNA 分析它们分属于细菌(广义的, Bacteria ,曾用 Eubacteria )、古菌 (Archaea ,曾用 Archaebacteria) 和 真核生物 (Eukaryota)( 图 1 · 1) 。非细胞生物包括病毒和亚病毒。 图 1 · 1 细胞生物的系统发育树 ( Madigan et al., 2000) 原核生物与真核生物的区别 原核生物包括古菌和细菌,与真核生物的区 别综合列于表 1-1 。主要差异有( 1 )、原核生 物的遗传物质主要是以双螺旋 DNA 构成的一条染 色体 (chromosome) ,仅形成一个核区,没有核膜 包围,无核仁,称为原核 (nucleoid) 或拟核,无 组蛋白与之相结合。真核生物的遗传物质以双螺旋 DNA 构成一条或一条以上的多条染色体群,形成一 个真核 (nucleolus) ,有一核膜包围,膜上有孔, 有核仁,明显有别于周围的细胞质,并有组蛋白与 之相结合。而且各种细胞器如线粒体、叶绿体携带 有自己的 DNA ,可自主复制。( 2 )、原核生物 细胞的细胞质由细胞膜 (cell membrane) 包围, 并有细胞膜大量褶皱内陷入细胞质中形成中间体 或称为间体 (mesosome) 。不含其他分化明显的细 胞器 (organelles) 。真核生物细胞同样由细胞膜 包围,但不内陷,内含多种细胞器,如主要进行呼吸能量代谢的线粒体( mitochondria )和光合作用的叶绿体 (chloroplast) 等。各种细胞器有各自的膜包围,细胞器膜与细胞膜之间无直接关系。( 3 )、原核生物和真核生物 细胞的蛋白质合成都是在核蛋白体上进行,但大小不同,原核生物的核蛋白体为 70S ,而真核生物的核蛋白体为 80S , 其细胞器的核蛋白体也为 70S 。而且它们各自的亚单位构成也不一样,原核生物的核蛋白体是由 50S 和 30S 的两个 亚单位构成,真核生物的核蛋白体是由 60S 和 40S 两个亚单位构成,各亚单位的构成上也有区别。 古菌、细菌和真核生物三域 原核生物中,古菌和细菌在细胞形态结构、生长繁殖、生理代谢、遗传物质存在方式等方面相类似,因而同属原 核生物。但在分子生物学水平上,古菌和细菌之间有明显差别,主要表现见表 1-1 。从这些差异可见,古菌确是不仅 在细胞化学组成上更是在分子生物学水平上不同于同属于原核生物的细菌和真核生物的另一类特殊生物类群。 古菌 (archaea) 、细菌 (bacteria) 和真核生物 (eucaryoutes) 三域 (urkingdoms) 的概念是沃斯 (Woese) 及 其同事 1977 年根据对代表性细菌类群的 16S rRNA 碱基序列进行广泛比较后提出的,认为生物界的发育并不是一个 由简单的原核生物发育到较完全、较复杂的真核生物的过程,而是明显存在着三个发育不同的基因系统,即古菌、细 菌和真核生物。并认为这三个基因系统几乎是同时从某一起点各自发育而来,这一起点即是至今仍不明确的一个原始 祖先。这一生物界三域观念已被广泛接受。 虽然从系统发育来看,细菌和古菌是二种不同的生物类群,但它们的细胞形态和结构却基本一致,同属原核生物 (Procaryotes) ,原核生物是指一大类细胞不具核膜,也无核仁,只有核区,称为原核的单细胞生物。本章介绍原核 微生物的形态、结构和功能
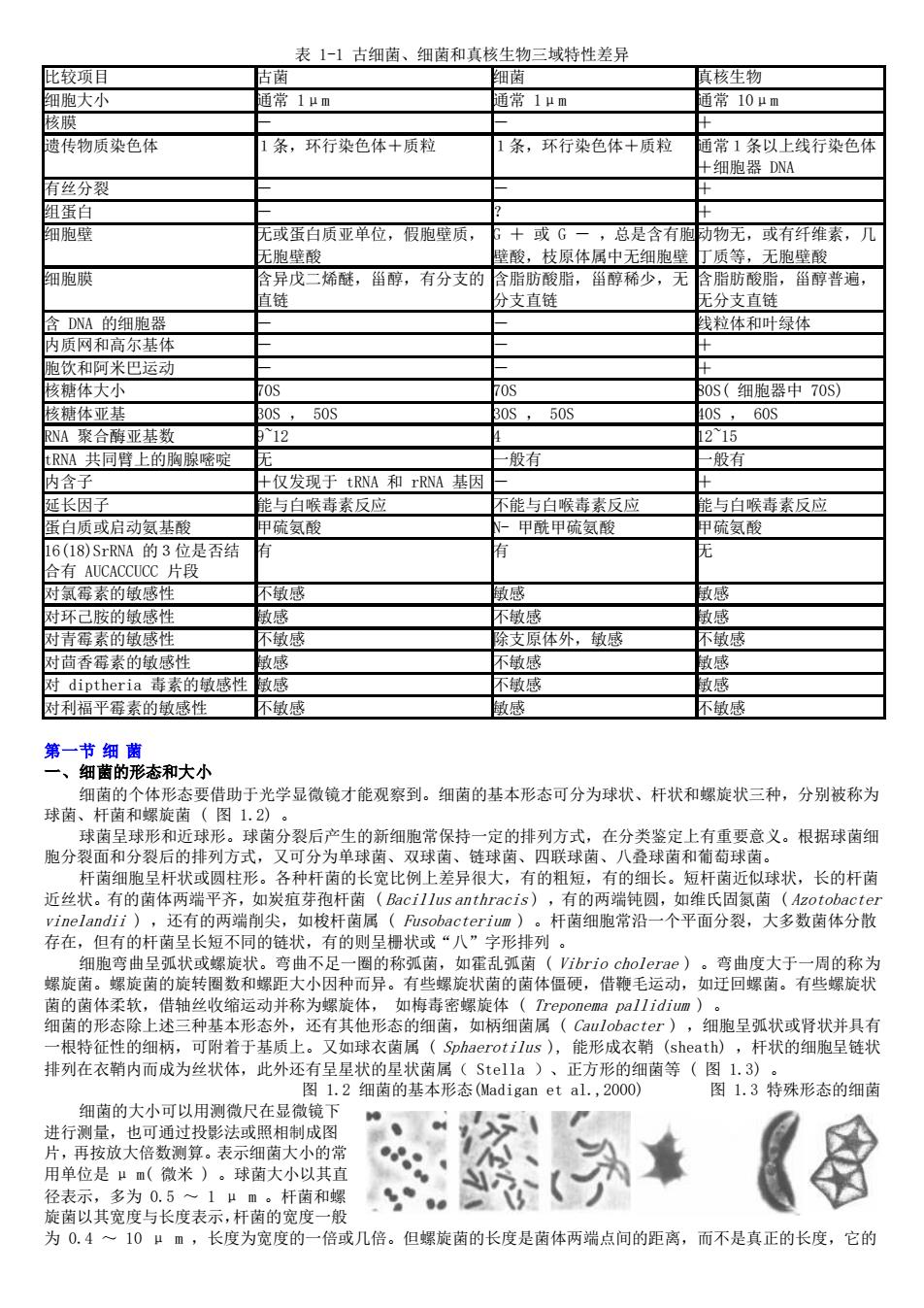
表1-1古细茵、细菌和真核生物三域特性差异 比较项目 古有 细 ]真核生物 细胞大小 常1山m 币常1山m 币常10um 核膜 遗传物质染色体 条,环行染色体十质粒 1条,环行染色体+质粒 通常1条以上线行染色体 +细胸器DWA 有丝分裂 组蛋白 细跑壁 或蛋白质亚单位,假胞壁质, 或G ,总是今有肠 幼物无,或有纤维素,几 无胞壁酸 庭酸,枝原体屈中无细胞壁 丁质等,无胞壁酸 细胞膜 含异戊二烯醚,甾醇,有分支的 含脂肪酸脂,留醇稀少,无 含脂肪酸脂,甾醇普遍, 直 分支直结 无分支直链 全DNA的细朐器 线行休和叶绿依 内质网和高尔基体 向饮知河米四云动 核镜体大小 0S(细胞哭中70S) 核纳休形甘 50 50S 0S.60S NA聚合酶亚其新 15 NA共同壁上的胸感啶 妈右 仅发现于tRNA和rRNA基因 白质或启动氨基酸 氨酸 毒素反应 能与白喉毒素反应 与白喉毒素反应 有C的片 的 背素 体外,敏感 对茴香莓素的敏感性 对diptheria毒素的敏感性敏感 利福平霉素的敏感性 不成 第一 态和 要借助于光学显微镜才能观察到。细菌的基本形态可分为球状、杆状和螺旋状三种,分别被称为 球菌 球菌县球形和球形 ,球菌分裂后产生的新细胞常保持一定的排列方式,在分类鉴定上有重要意义。根据球菌细 胞分裂面和分裂后的排列方式,又可分为单球菌、双球菌、链球菌、四联球菌、八叠球菌和葡萄球菌。 杆菌细胞呈杆状或圆柱形。各种杆菌的长宽比例上差异很大,有的粗短,有的细长。短杆南近似球状,长的杆菌 近丝状。有的菌体两端平齐,如炭疽芽孢杆菌(Bacillus anthracis),有的两端纯圆,如维氏固氮菌(Azotobacter vinelandii),还有的两端削尖,如梭杆菌属(Fusobacterium)。杆菌细胞常沿一个平面分裂,大多数菌体分散 存在,但有的杆菌呈长短不同的链状,有的则呈橱状或“八”字形排列。 菌的旋转圈 小因种而异· 并称为螺 击 状 上述二种基本形 不有其形态的细菌, 细胞呈弧状或肾状并具有 Oius),能形成衣鞘(sheath),杆状的细胞呈链状 排列在衣箱内而成为丝状体,此外还有呈星状的星状菌属(St11a)、正方形的细菌等(图3》 图1.2细菌的基本形态(ladigan et al.,2000) 图1.3特殊形态的细菌 细菌的大小可以用测微尺在显微镜下 进行测量,也可通过投影法或照相制成图 片,再按放大倍数测算。表示细菌大小的常 用单位是“m(微米)。球茵大小以其直 《 1μm。杆和螺 为0.4一10μm,长度为宽度的一倍或几倍。但螺旋菌的长度是菌体两端点间的距离,而不是其正的长度,它的
表 1-1 古细菌、细菌和真核生物三域特性差异 比较项目 古菌 细菌 真核生物 细胞大小 通常 1μm 通常 1μm 通常 10μm 核膜 - - + 遗传物质染色体 1条,环行染色体+质粒 1条,环行染色体+质粒 通常1条以上线行染色体 +细胞器 DNA 有丝分裂 - - + 组蛋白 - ? + 细胞壁 无或蛋白质亚单位,假胞壁质, 无胞壁酸 G + 或 G - ,总是含有胞 壁酸,枝原体属中无细胞壁 动物无,或有纤维素,几 丁质等,无胞壁酸 细胞膜 含异戊二烯醚,甾醇,有分支的 直链 含脂肪酸脂,甾醇稀少,无 分支直链 含脂肪酸脂,甾醇普遍, 无分支直链 含 DNA 的细胞器 - - 线粒体和叶绿体 内质网和高尔基体 - - + 胞饮和阿米巴运动 - - + 核糖体大小 70S 70S 80S( 细胞器中 70S) 核糖体亚基 30S , 50S 30S , 50S 40S , 60S RNA 聚合酶亚基数 9~12 4 12~15 tRNA 共同臂上的胸腺嘧啶 无 一般有 一般有 内含子 +仅发现于 tRNA 和 rRNA 基因 - + 延长因子 能与白喉毒素反应 不能与白喉毒素反应 能与白喉毒素反应 蛋白质或启动氨基酸 甲硫氨酸 N- 甲酰甲硫氨酸 甲硫氨酸 16(18)SrRNA 的3位是否结 合有 AUCACCUCC 片段 有 有 无 对氯霉素的敏感性 不敏感 敏感 敏感 对环己胺的敏感性 敏感 不敏感 敏感 对青霉素的敏感性 不敏感 除支原体外,敏感 不敏感 对茴香霉素的敏感性 敏感 不敏感 敏感 对 diptheria 毒素的敏感性 敏感 不敏感 敏感 对利福平霉素的敏感性 不敏感 敏感 不敏感 第一节 细 菌 一、细菌的形态和大小 细菌的个体形态要借助于光学显微镜才能观察到。细菌的基本形态可分为球状、杆状和螺旋状三种,分别被称为 球菌、杆菌和螺旋菌 ( 图 1.2) 。 球菌呈球形和近球形。球菌分裂后产生的新细胞常保持一定的排列方式,在分类鉴定上有重要意义。根据球菌细 胞分裂面和分裂后的排列方式,又可分为单球菌、双球菌、链球菌、四联球菌、八叠球菌和葡萄球菌。 杆菌细胞呈杆状或圆柱形。各种杆菌的长宽比例上差异很大,有的粗短,有的细长。短杆菌近似球状,长的杆菌 近丝状。有的菌体两端平齐,如炭疽芽孢杆菌 ( Bacillus anthracis ) ,有的两端钝圆,如维氏固氮菌 ( Azotobacter vinelandii ) ,还有的两端削尖,如梭杆菌属 ( Fusobacterium ) 。杆菌细胞常沿一个平面分裂,大多数菌体分散 存在,但有的杆菌呈长短不同的链状,有的则呈栅状或“八”字形排列 。 细胞弯曲呈弧状或螺旋状。弯曲不足一圈的称弧菌,如霍乱弧菌 ( Vibrio cholerae ) 。弯曲度大于一周的称为 螺旋菌。螺旋菌的旋转圈数和螺距大小因种而异。有些螺旋状菌的菌体僵硬,借鞭毛运动,如迂回螺菌。有些螺旋状 菌的菌体柔软,借轴丝收缩运动并称为螺旋体, 如梅毒密螺旋体 ( Treponema pallidium ) 。 细菌的形态除上述三种基本形态外,还有其他形态的细菌,如柄细菌属 ( Caulobacter ) ,细胞呈弧状或肾状并具有 一根特征性的细柄,可附着于基质上。又如球衣菌属 ( Sphaerotilus ), 能形成衣鞘 (sheath) ,杆状的细胞呈链状 排列在衣鞘内而成为丝状体,此外还有呈星状的星状菌属( Stella )、正方形的细菌等 ( 图 1.3) 。 图 1.2 细菌的基本形态(Madigan et al.,2000) 图 1.3 特殊形态的细菌 细菌的大小可以用测微尺在显微镜下 进行测量,也可通过投影法或照相制成图 片,再按放大倍数测算。表示细菌大小的常 用单位是 μ m( 微米 ) 。球菌大小以其直 径表示,多为 0.5 ~ 1 μ m 。杆菌和螺 旋菌以其宽度与长度表示,杆菌的宽度一般 为 0.4 ~ 10 μ m ,长度为宽度的一倍或几倍。但螺旋菌的长度是菌体两端点间的距离,而不是真正的长度,它的
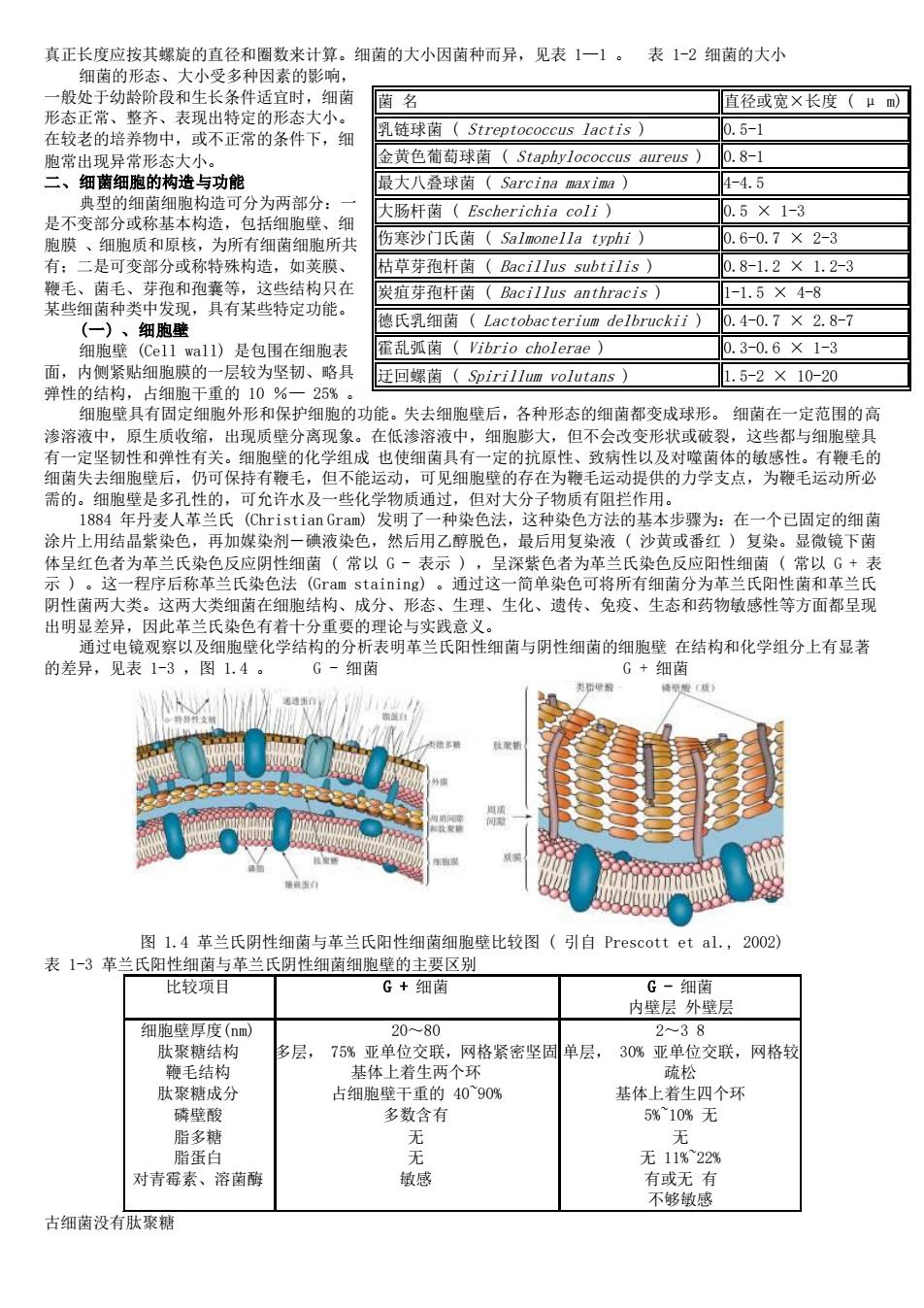
真正长度应按其螺旋的直径和圈数来计算。细菌的大小因菌种而异,见表1一1。 表1-2细菌的大小 菌名 直径或宽X长度(um) 在老的培养物中,或不正常的条件下】 乳链球菌(Streptococcus lactis 金黄色葡萄球黄(Staphvlococcus aureus) 0.8-1 最大八叠球菌(Sarcina maxim) 1-4.5 典型的细菌细胞构造可分为两部分: 大肠杆菌(Escherichia coli) 0.5×1-3 是不变部分或称基本构造,包括细胞壁、细 胞膜、细胞质和原核,为所有细菌细胞所共 伤寒沙门氏菌(Salmonella typhi) 0.6-0.7×2-3 有:二是可变部分或称特殊构造,如荚膜、 枯草芽孢杆菌(Bacil1 us subtilis) 0.8-1.2×1.2-3 这些结构只在 炭疽芽孢杆菌(Bacillus anthracis】 1-1.5×4-8 某些细菌种头 德氏细菌(Lactobacterium delbruckii) 0.4-0.7×2.8-7 a11)是包 乱弧南(Vibrio cholerae) 12-06×1-3 回螺(Spirillum volutans)】 1.5-2×10-20 细胞壁只有固定细胞外形和保护细胞的功能。失去细胞壁后,各种形态的细菌都变成球形。 细黄在一定带用的 渗溶液中,原生质收缩,出现质壁分离现象。在低渗溶液中,细胞膨大,但不会改变形状或破裂,这些都与细跑壁男 有一定坚韧性和弹性有关。细胞壁的化学组成也使细菌具有一定的抗原性、致病性以及对噬茵体的敏感性。有鞭毛的 细菌失去细胞壁后,仍可保持有鞭毛,但不能运动,可见细胞壁的存在为鞭毛运动提供的力学支点,为鞭毛运动所必 需的。细胞壁是多孔性的,可允许水及一些化学物质通过,但对大分子物质有阻拦作用。 L884 、革兰氏 法的举 为:在 个已固定的细菌 涂片 结晶 再加媒染 碘液染 发后用2种· 黄或 复染。 生细菌(常以i 表示 这两大细黄 在细胞结 发示通过这单速色可将所有细菌分为革兰民阳西品风 分形态生理、生化、遗传、免疫 出明显差异,因此革兰氏染色有着十分重要的理论与实践意义 通过电镜观察以及细胞壁化学结构的分析表明革兰氏阳性细菌与阴性细菌的细胞壁在结构和化学组分上有显著 的差异,见表1-3,图1.4。 G-细茵 G+细茵 图1.4革兰氏阴性细菌与革兰氏阳性细菌细胞壁比较图(引自Prescott et al.,2002) 表1-3革兰氏阳性细菌与革兰 氏阴性细菌细胞壁的主要区别 比较项 G+图 G二细菌 内壁层外壁层 38 层, 网格紧密坚同 单层, 30%亚单 位交联 网格书 肽聚糖成 4090% 多数含有 脂蛋白 无11%22% 对青莓素、溶菌酶 敏感 有或无有 不够敏成 古细菌设有肽聚糖
真正长度应按其螺旋的直径和圈数来计算。细菌的大小因菌种而异,见表 1—1 。 表 1-2 细菌的大小 细菌的形态、大小受多种因素的影响, 一般处于幼龄阶段和生长条件适宜时,细菌 形态正常、整齐、表现出特定的形态大小。 在较老的培养物中,或不正常的条件下,细 胞常出现异常形态大小。 二、细菌细胞的构造与功能 典型的细菌细胞构造可分为两部分:一 是不变部分或称基本构造,包括细胞壁、细 胞膜 、细胞质和原核,为所有细菌细胞所共 有;二是可变部分或称特殊构造,如荚膜、 鞭毛、菌毛、芽孢和孢囊等,这些结构只在 某些细菌种类中发现,具有某些特定功能。 (一) 、细胞壁 细胞壁 (Cell wall) 是包围在细胞表 面,内侧紧贴细胞膜的一层较为坚韧、略具 弹性的结构,占细胞干重的 10 %— 25% 。 细胞壁具有固定细胞外形和保护细胞的功能。失去细胞壁后,各种形态的细菌都变成球形。 细菌在一定范围的高 渗溶液中,原生质收缩,出现质壁分离现象。在低渗溶液中,细胞膨大,但不会改变形状或破裂,这些都与细胞壁具 有一定坚韧性和弹性有关。细胞壁的化学组成 也使细菌具有一定的抗原性、致病性以及对噬菌体的敏感性。有鞭毛的 细菌失去细胞壁后,仍可保持有鞭毛,但不能运动,可见细胞壁的存在为鞭毛运动提供的力学支点,为鞭毛运动所必 需的。细胞壁是多孔性的,可允许水及一些化学物质通过,但对大分子物质有阻拦作用。 1884 年丹麦人革兰氏 (Christian Gram) 发明了一种染色法,这种染色方法的基本步骤为:在一个已固定的细菌 涂片上用结晶紫染色,再加媒染剂-碘液染色,然后用乙醇脱色,最后用复染液 ( 沙黄或番红 ) 复染。显微镜下菌 体呈红色者为革兰氏染色反应阴性细菌 ( 常以 G - 表示 ) ,呈深紫色者为革兰氏染色反应阳性细菌 ( 常以 G + 表 示 ) 。这一程序后称革兰氏染色法 (Gram staining) 。通过这一简单染色可将所有细菌分为革兰氏阳性菌和革兰氏 阴性菌两大类。这两大类细菌在细胞结构、成分、形态、生理、生化、遗传、免疫、生态和药物敏感性等方面都呈现 出明显差异,因此革兰氏染色有着十分重要的理论与实践意义。 通过电镜观察以及细胞壁化学结构的分析表明革兰氏阳性细菌与阴性细菌的细胞壁 在结构和化学组分上有显著 的差异,见表 1-3 ,图 1.4 。 G - 细菌 G + 细菌 图 1.4 革兰氏阴性细菌与革兰氏阳性细菌细胞壁比较图 ( 引自 Prescott et al., 2002) 表 1-3 革兰氏阳性细菌与革兰氏阴性细菌细胞壁的主要区别 比较项目 G + 细菌 G - 细菌 内壁层 外壁层 细胞壁厚度(nm) 肽聚糖结构 鞭毛结构 肽聚糖成分 磷壁酸 脂多糖 脂蛋白 对青霉素、溶菌酶 20~80 多层, 75% 亚单位交联,网格紧密坚固 基体上着生两个环 占细胞壁干重的 40~90% 多数含有 无 无 敏感 2~3 8 单层, 30% 亚单位交联,网格较 疏松 基体上着生四个环 5%~10% 无 无 无 11%~22% 有或无 有 不够敏感 古细菌没有肽聚糖 菌 名 直径或宽×长度 ( μ m) 乳链球菌 ( Streptococcus lactis ) 0.5-1 金黄色葡萄球菌 ( Staphylococcus aureus ) 0.8-1 最大八叠球菌 ( Sarcina maxima ) 4-4.5 大肠杆菌 ( Escherichia coli ) 0.5 × 1-3 伤寒沙门氏菌 ( Salmonella typhi ) 0.6-0.7 × 2-3 枯草芽孢杆菌 ( Bacillus subtilis ) 0.8-1.2 × 1.2-3 炭疽芽孢杆菌 ( Bacillus anthracis ) 1-1.5 × 4-8 德氏乳细菌 ( Lactobacterium delbruckii ) 0.4-0.7 × 2.8-7 霍乱弧菌 ( Vibrio cholerae ) 0.3-0.6 × 1-3 迂回螺菌 ( Spirillum volutans ) 1.5-2 × 10-20

1、革兰氏阳性细菌细胞壁 氏阳性细菌只有一层厚约20~8Onm的细胞壁。细胞壁的化学组成以肤聚糖(Peptidoglycan)为主,占细 胞壁物质总量的40%~ 学成和码o1caci0又称酸,是+细菌细能特有的成分 肽聚糖是除古菌外 N-乙酰壁酸(M 多层网状结焚是中 有 。肽聚糖的单体含有 )聚合而成 CH:OH 种组 乙酰葡萄糖胺N-acetylglu 乙 酰胞壁酸(N-acetylmuramic acid,简称NAM)和四肽链。 N 乙酰葡萄糖胺与N乙酰胞壁酸交替排列,通过B-1,4糖 苷健连接成聚糖链骨架。四肽链则是通过 一个酰胺健与N乙酰 HC-CH CH, 溶能酶装感健 胞壁酸相连,肽聚糖单体聚合成肽聚糖大分子时,主要是两条不 同聚糖链骨架上与N 乙酰胞壁酸相连的两条相邻四肽链间的 HC-CHO L-丙氢形 父 m 不的组 四肽链 谷 CH-CO NH; D 1 少数是甘氨酸或 丝氢酸。而的变化较大,可以是内消旋的二氨基庚二酸 m -DAP) I-DAP 鸟氨酸, 氨基丁酸,有时也可以是同型丝氨酸或L-丙氨酸。四肽链第二位的D-谷氨酸也可羟基化,游离的▣一羟基可酰胺 化或被甘氨酸等所取代。革兰氏阳性菌(以金黄色葡萄球菌为例)的四肽链是L丙氨酸-D谷氨酸L-赖氨酸 D-内氨酸, 二条四肽链间通过五聚甘氨酸桥肽链而间接交联:桥肽的一头连接L一赖氨酸的€ 氨基,另一头连 着另 条四肽链的D丙氨酸的羟基,交联度高,从而形成了紧密编织、质地坚硬和机械性强度很大的多层三维空间 网格结 图1.6肽聚糖单层结构模式图(引自Prescott et al.,2002) A.革兰氏阴性闲 B.革兰氏阳性 壁质酸是大多数革兰氏阳性菌细 -NAM-NAG- -NAG 干重的50% L-A 酰壁酸相结 D-Gl D-A 合。此酸有两个类型:甘油型磷壁质酸 DAP 0-G D-Ala DAP L-Lys (图1.7)和核醇型磷壁质酸。甘油型磷 0-Ala 壁质酸是由许多分子的甘油借磷酸二酯键 联结起来的分子:核醇型磷壁质酸是由君 -NAG NAG 设认 的完整性 国1.7瞬壁酸类型及基本结构 核糖醇磷壁酸B。甘油型磷壁酸 内壁层紧贴细胞膜,厚约2~3m,由肽聚糖组成,占细 胞壁干重的5%~10%。外壁层又称外膜(outer membrane) /m)iH 约8~10nm,主要由脂多糖(1 ipopolysaccharide,LPS)和 (EI BI)-H 外膜蛋白(out membrane proteins)组成。 ,菌与革兰氏阳性菌肽聚糖的不同之处就在于它们短 H.L-I: 肤上 吸以及两承湿 性菌〈以大肠杆南为的 进 个氨基酸是 谷氨酸 条肽链联结起来 脂多糖是G-茵细胞壁的特有成分,G+茵中不存在。脂多糖由三部分组成,即0-侧链、核心多糖和类脂A(图 .8)。0-侧结向外 由若干个低案糖的重复单位组成,由于具有抗原性,故又称0一抗原或茵体抗原。不同种或型的细 菌,0侧链的组成和结构(如多糖的种类和序列)均有变化,构成了各自的特异性抗原。像沙门氏菌(Sa101a), 根据0-抗原可再细分为1000多个血清型,这些血清型的沙门氏菌,核心多糖部分相同,而0-抗原的差异使之在 免疫学和临床诊断中具有重要意义。非致病性革兰氏阴性细菌细胞壁组成中不具0侧链。核心多糖由庚糖、半乳糖 2酮基-3-脱氧辛酸组成,所有革兰氏阴性细菌都有此结构。类脂A是以脂化的葡萄糖胺二糖为单位,通过焦磷酸 键组成的一种独特的糖脂化合物。类脂A的结构在不同细菌中有所不同,它是革兰氏阴性细菌内毒素的毒性中心。 外膜蛋白指嵌合在脂多糖和磷脂层外膜上的20多种蛋白,多数功能还不清楚。其中脂蛋白(lipoprotein)的
1、革兰氏阳性细菌细胞壁 革兰氏阳性细菌只有一层厚约 20 ~ 80nm 的细胞壁。细胞壁的化学组成以肽聚糖 (Peptidoglycan) 为主,占细 胞壁物质总量的 40 %~ 90% 。另外还结合有磷壁质酸 (teichoic acid) 又称垣酸,是 G + 细菌细胞壁特有的成分。 图 1.5 肽聚糖的化学组成和一级结构 肽聚糖 是除古菌外,凡有细胞壁的原核生物细胞壁中的共 有组分。肽聚糖是由若干肽聚糖单体 ( 图 1. 5 ) 聚合而成的 多层网状结构大分子化合物。肽聚糖的单体含有三种组分: N- 乙酰葡萄糖胺 (N-acetylgluco samine ,简写 NAG) 、 N- 乙 酰胞壁酸 (N-acetylmuramic acid ,简称 NAM) 和四肽链。 N- 乙酰葡萄糖胺与 N- 乙酰胞壁酸交替排列,通过 β -1 , 4 糖 苷键连接成聚糖链骨架。四肽链则是通过一个酰胺键与 N- 乙酰 胞壁酸相连,肽聚糖单体聚合成肽聚糖大分子时,主要是两条不 同聚糖链骨架上与 N- 乙酰胞壁酸相连的两条相邻四肽链间的 相互交联 ( 见图 1.6 ) 。不同种类细菌的肽聚糖聚糖链骨架基 本是相同的,不同的是四肽链氨基酸的组成以及两条四肽链间的 交联方式。四肽链一般可以用 R 1 -D- 谷氨酸 -R 3 -D- 丙氨 酸的通式表示。 R 1 大多是 L- 丙 氨酸,少数是甘氨酸或 L- 丝氨酸。而 R3 的变化较大,可以是内消旋的二氨基庚二酸 (meso-DAP) , L- 赖氨酸, L , L-DAP , L- 鸟氨酸, L- 二 氨基丁酸,有时也可以是同型丝氨酸或 L- 丙氨酸。四肽链第二位的 D- 谷氨酸也可羟基化,游离的 α - 羟基可酰胺 化或被甘氨酸等所取代。革兰氏阳性菌 ( 以金黄色葡萄球菌为例 ) 的四肽链是 L- 丙氨酸 -D- 谷氨酸 -L- 赖氨酸 -D- 丙氨酸,二条四肽链间通过五聚甘氨酸桥肽链而间接交联;桥肽的一头连接 L- 赖氨酸的 ε - 氨基,另一头连接 着另一条四肽链的 D- 丙氨酸的羟基,交联度高,从而形成了紧密编织、质地坚硬和机械性强度很大的多层三维空间 网格结构。 图 1.6 肽聚糖单层结构模式图 ( 引自 Prescott et al., 2002) A. 革兰氏阴性菌 B. 革兰氏阳性 菌 磷壁质酸是大多数革兰氏阳性菌细胞 壁组分,占细胞壁干重的 50% 左右,以磷 酸二酯键同肽聚糖的 N- 乙酰胞壁酸相结 合。此酸有两个类型:甘油型磷壁质酸 ( 图 1.7) 和核醇型磷壁质酸。甘油型磷 壁质酸是由许多分子的甘油借磷酸二酯键 联结起来的分子;核醇型磷壁质酸是由若 干分子的核醇借磷酸二酯键联结而成的分 子。一般认为磷壁酸因含有大量的带负电 性的磷酸,故大大加强了细胞膜对二价离子的吸附,尤其是镁离子。而高浓度的镁离子有利于对维持细胞膜的完整性、 提高细胞壁合成酶的活性。磷壁酸是革兰氏阳性菌表面抗原 (C 抗原 ) 的主要成分,也是噬菌体吸附的受体位点。 图 1.7 磷壁酸类型及基本结构 A. 核糖醇磷壁酸 B. 甘油型磷壁酸 2、革兰氏阴性菌细胞壁 G - 菌的细胞壁比 G + 菌的薄,可分为内壁层和外壁层。 内壁层紧贴细胞膜,厚约 2 ~ 3nm ,由肽聚糖组成,占细 胞壁干重的 5%~10%。外壁层又称外膜(outer membrane) 约 8~10nm,主要由脂多糖 (lipopolysaccharide,LPS) 和 外膜蛋白 (out membrane proteins) 组成。 G - 菌与革兰氏阳性菌肽聚糖的不同之处就在于它们短 肽上的氨基酸以及两条短肽上氨基酸相联结的方式不同。革 兰氏阴性菌 ( 以大肠杆菌为例 ) 肽聚糖肽链中的四个氨基酸是 L- 丙氨酸、 D- 谷氨酸、内消旋二氨基庚二酸及 D- 丙氨酸。一股肽链第三位上的二氨基庚二酸的游离氨基与相邻的另一股肽链末端的 D- 丙氨酸的羧基形成肽键,将两 条肽链联结起来。 脂多糖是 G - 菌细胞壁的特有成分,G + 菌中不存在。脂多糖由三部分组成,即 O- 侧链、核心多糖和类脂 A( 图 1.8) 。O-侧链向外,由若干个低聚糖的重复单位组成,由于具有抗原性,故又称 O-抗原或菌体抗原。不同种或型的细 菌,O-侧链的组成和结构 ( 如多糖的种类和序列 ) 均有变化,构成了各自的特异性抗原。像沙门氏菌( Salmonella ), 根据 O- 抗原可再细分为 1000 多个血清型,这些血清型的沙门氏菌,核心多糖部分相同,而 O- 抗原的差异使之在 免疫学和临床诊断中具有重要意义。非致病性革兰氏阴性细菌细胞壁组成中不具 O- 侧链。核心多糖由庚糖、半乳糖、 2- 酮基 -3- 脱氧辛酸组成,所有革兰氏阴性细菌都有此结构。类脂 A 是以脂化的葡萄糖胺二糖为单位,通过焦磷酸 键组成的一种独特的糖脂化合物。类脂 A 的结构在不同细菌中有所不同,它是革兰氏阴性细菌内毒素的毒性中心。 外膜蛋白指嵌合在脂多糖和磷脂层外膜上的 20 多种蛋白,多数功能还不清楚。其中 脂蛋白( lipoprotein ) 的

蛋白质部分末端游离的氨基酸残基与肽聚糖层的某些二氨基庚二酸残基形成肽键,呈共价结合,其脂质部分同外壁层 脂相结合:因 糖层到所 通道子的亲水性物质得以进出。装血 的蛋白存在于G 的白形成“充水 一些孔蛋白具有 度特异性 种或多种物质的特异性结合位点。最大的微孔蛋白可以允许分子量高达50D的物质进 图1,8革兰氏G-细菌脂多糖、 类脂 、砖脂 、孔蛋白的排列方式 3、细胞壁结构与菲兰氏垫色的关系 革兰氏染色的性质同细胞壁的结构与组 分省关。在大多认为。在垫伍程中。细 胞内形成了一种不溶性的结品紫-碘的复合 物,这种复合物可被乙醇(或丙酮)从G细茵 细胞内抽提出来,但不能从G菌中抽提出来, 区是田 脂质含量低 细胞壁较厚,肽聚糖含量高 品紫-碘复 是保留初沈的生 而革兰氏阴性细菌细胞壁肽聚糖层较薄 含量较少,而且脂质含量高,经乙醇处理后 脂质被溶解,渗透性增高,结果结品紫碘复 合物外渗,细胞被番红复染成红色。 4、细胞壁缺陷型细菌 用溶菌酶处理细胞或在培养基中加入青霉素、甘氨酸或丝裂霉素C等因子,便可破坏或抑制细胞壁的形成,成为 细胞壁缺陷细菌 质体 原生质球和细菌L-型。用溶菌酶除去革兰氏阴性茵细胞壁时,若先用乙二胺 四乙 迪过 而且特别脆弱。 离以至通气等因素都易引起其破裂 有的原生质体还保目 但不能 不能被相应的噬茵体感染。 原生质体在适宜条件下同样可生长繁殖 其他生物活本 如用即将形 成芽孢的营养体获得的原生质体仍可形成芽孢。原生质体的获得,给微生物学工作者提供了另一种类型的生物学实体, 用原生质体融合新技术,可培有新的优良菌种。 原生质球(Spheroplast)指细胞壁未被全部去掉的细菌细胞,它呈圆球状,可以人为地通过溶菌酶或青霉素处理 革兰氏阴性细菌而获得。该类菌细胞壁肽聚糖虽被除去,但外壁层中脂多糖、脂蛋白仍然保留,外壁的结构尚存。所 以,原生质球较之原生质体对外界环境具一定抗性, 细菌L-型(bacterial上for 环境条件下因基因突变而产生的无壁类型。细胞呈多形态,有的 能通过细商 比是能 定”变异株。由于它最先被英国Liste 医 究院发现 故名细门 间,指位于细胞壁与细胞质膜之间的狭小空间 对细菌的营养吸收、楼酸代谢、趋化性和抗药性等常有直要作用。质外膨的种类和数量随菌种而异,目 细菌)中发现的质外酶主要有:RA酶I、DNA内切酶I、青霉素酶及许多磷酸化酶等。 (二)、细胞质膜 细胞质膜(cytoplasmic membrane))又称细胞膜(cell membrane),是围绕在细胞质外面的一层柔软而富有弹性 的薄膜,厚约8m。细菌细胞膜占细胞干重的10%左右,其化学成分主要为脂类(20%30%)与蛋白质(60%70%)。 原核生物中除支原体外,细胞膜上一般不含胆固醇,这与真核生物不同(图1.9) 图1.9原核生物与真核生物的细胞质膜比较 细菌细胞膜的脂类主要为甘 由磷脂 磷脂分子在 风分子层的外表面 疏水的非极性基朝内(即排列在组成膜的内侧 面),这样就形成了膜的基本骨架。磷脂中的脂肪酸 有饱和及不饱和两种,膜的流动性高低主要取决于它 们的相对含量和类型,如低温型微生物的膜中含有较 多的不饱和脂肪酸,而高温型微生物的膜则富含饱和 脂肪酸,从而保持了膜在不同温度下的正常生理功能。 细胞膜中的蛋白质依其存在位置可分为外周蛋白 。外周蛋白存在于膜的内或外表面 系水溶性蛋白
蛋白质部分末端游离的氨基酸残基与肽聚糖层的某些二氨基庚二酸残基形成肽键,呈共价结合,其脂质部分同外壁层 磷脂相结合。因此,脂蛋白是从肽聚糖层到外壁层之间的桥梁。另有一类称 微 孔蛋白( porin ) 的蛋白存在于 G - 菌的外壁层中,这些蛋白的功能是作为一个通道使低分子的亲水性物质得以进出。有特异性性与非特异性两类。特异 性孔蛋白形成“充水”的通道,任何类型的小物质都可以通过。而另一些孔蛋白具有高度特异性,是因为它们含有一 种或多种物质的特异性结合位点。最大的微孔蛋白可以允许分子量高达 5000Da 的物质进入。 图 1.8 革兰氏 G - 细菌脂多糖、类脂 A 、磷脂、孔蛋白的排列方式 3 、细胞壁结构与革兰氏染色的关系 革兰氏染色的性质同细胞壁的结构与组 分有关。现在大多认为,在染色过程中,细 胞内形成了一种不溶性的结晶紫-碘的复合 物,这种复合物可被乙醇(或丙酮)从 G -细菌 细胞内抽提出来,但不能从 G +菌中抽提出来。 这是由于 G +菌细胞壁较厚,肽聚糖含量高, 脂质含量低甚或没有,经乙醇处理后引起脱 水,结果肽聚糖孔径变小,渗透性降低,结 晶紫-碘复合物不能外流,于是保留初染的紫 色。而革兰氏阴性细菌细胞壁肽聚糖层较薄, 含量较少,而且脂质含量高,经乙醇处理后, 脂质被溶解,渗透性增高,结果结晶紫-碘复 合物外渗,细胞被番红复染成红色。 4 、细胞壁缺陷型细菌 用溶菌酶处理细胞或在培养基中加入青霉素、甘氨酸或丝裂霉素 C 等因子,便可破坏或抑制细胞壁的形成,成为 细胞壁缺陷细菌,通常包括原生质体、原生质球和细菌 L- 型。用溶菌酶除去革兰氏阴性菌细胞壁时,若先用乙二胺 四乙酸 (EDTA) 处理外壁,则效果更好。 原生质体 (protoplast) 在革兰氏阳性细菌培养物中加溶菌酶或通过青霉素阻止其细胞壁的正常合成而获得的完 全缺壁细胞称原生质体。原生质体由于没有坚韧的细胞壁,故任何形态的菌体均呈球形。原生质体对环境条件很敏感, 而且特别脆弱,渗透压、振荡、离心以至通气等因素都易引起其破裂。有的原生质体还保留着鞭毛,但不能运动,也 不能被相应的噬菌体感染。原生质体在适宜条件下同样可生长繁殖,形成菌落,其他生物活性基本不变。如用即将形 成芽孢的营养体获得的原生质体仍可形成芽孢。原生质体的获得,给微生物学工作者提供了另一种类型的生物学实体, 用原生质体融合新技术,可培育新的优良菌种。 原生质球 (Spheroplast) 指细胞壁未被全部去掉的细菌细胞,它呈圆球状,可以人为地通过溶菌酶或青霉素处理 革兰氏阴性细菌而获得。该类菌细胞壁肽聚糖虽被除去,但外壁层中脂多糖、脂蛋白仍然保留,外壁的结构尚存。所 以,原生质球较之原生质体对外界环境具一定抗性,并能在普通培养基上生长。 细菌 L- 型 (bacterial L-form) 是细菌在某些环境条件下因基因突变而产生的无壁类型。细胞呈多形态,有的 能通过细菌滤器,故又称“滤过型菌”。 L- 型菌落生长缓慢,一般需经 2-7 天方见到针尖样小菌落,中心部分深埋 于培养基内,呈典型的“油煎蛋”状。这些变异型,有些是能回复至亲代的“不稳定”变异株,有些是不能回复的“稳 定”变异株。由于它最先被英国 Lister 医学研究院发现,故名细菌 L- 型。 周质空间 (Periplasmic space) 又称壁膜空间,指位于细胞壁与细胞质膜之间的狭小空间。内含质外酶。质外酶 对细菌的营养吸收、核酸代谢、趋化性和抗药性等常有重要作用。质外酶的种类和数量随菌种而异,目前已在细菌 ( 尤 其是 G - 细菌 ) 中发现的质外酶主要有: RNA 酶 I 、 DNA 内切酶 I 、青霉素酶及许多磷酸化酶等。 (二) 、细胞质膜 细胞质膜 (cytoplasmic membrane) 又称细胞膜 (cell membrane) ,是围绕在细胞质外面的一层柔软而富有弹性 的薄膜,厚约 8nm 。细菌细胞膜占细胞干重的 10% 左右,其化学成分主要为脂类 (20%~30%) 与蛋白质 (60%~70%) 。 原核生物中除支原体外,细胞膜上一般不含胆固醇,这与真核生物不同 ( 图 1.9) 。 图 1.9 原核生物与真核生物的细胞质膜比较 细菌细胞膜的脂类主要为甘油磷脂。磷脂分子在 水溶液中很容易形成具有高度定向性的双分子层,相 互平行排列,亲水的极性基指向双分子层的外表面, 疏水的非极性基朝内 ( 即排列在组成膜的内侧 面 ) ,这样就形成了膜的基本骨架。磷脂中的脂肪酸 有饱和及不饱和两种,膜的流动性高低主要取决于它 们的相对含量和类型,如低温型微生物的膜中含有较 多的不饱和脂肪酸,而高温型微生物的膜则富含饱和 脂肪酸,从而保持了膜在不同温度下的正常生理功能。 细胞膜中的蛋白质依其存在位置可分为外周蛋白和内 嵌蛋白两大类。外周蛋白存在于膜的内或外表面,系水溶性蛋白,占膜蛋白总量的 20 %~ 30% 。内嵌蛋白又称固有 蛋白或结构蛋白,镶嵌于磷脂双层中,多为非水溶性蛋白,占总量的 70 %~ 80%( 图 1.10) 。膜蛋白除作为膜的结