酵母线粒体、原生动物纤毛虫也有类似情形。所以结论应该是:遗传密码并非是绝对 通用的,而是近于完全通用的。 表12-6人线粒体DWA中密码编制特点 通常情汉下可绝巴的 T线粒体DNA所编码的 终止信号 色 色氨 色钉 导意氧破 甲砖氨酸 AUG 甲硫氨酸 甲硫氨酸 AGA 精氨酸 终止信号 结氨酸 终止信号 第二节 蛋白质合成的分子基础 氨基酸是在核糖体上加入到多肽链中的。在与mRNA作用之前,氨基酸先共价地与转 运RNA((transfer RNA,tRNA)形成氨酰tRNA。氨酰-tRNA结合到mRNA的特殊位点上。 mRNA含有遗传密码的信息,用于指导特定氨基酸序列多肽链的合成。一个核糖体结合到 一个mRNA分子合成起始序列上,并由此开始读码,沿着密码序列合成一条多肽链。读码 的方向是从mRNA的5'到3',而合成出来的多肽则是从氨基端到羧基端。通常, 一个 mRNA分子上可结合有多个不同时间开始翻译的核糖体,这样的结构称为多聚核糖体 (polysome)。在原核细胞中,mRNA的转录与多肽的翻译是同时进行的,如图12-1所示,可 见在染色体DNA分子上,有多条正在转录的mRNA分子,每个分子上都结合有多个正在进 行翻译的核糖体。真核生物的转录与翻译是在不同的地方进行的,核糖体可以自由地存在 于细胞质中,或者与内质网膜结合。RNA分子在蛋白质的合成中起若举足轻重的作用,除 了mRNA和tRNA,核糖体还含有核糖体RNA(ribosome RNA,RNA)。 mRNA是蛋白质合成的模板 JacobF.和MonodJ.早在1961年就已提出mRNA的概念。他们认为,既然蛋白质是在细胞质 中合成的,而编码蛋白质的信息载体DNA却在细胞核内,所以必定有一种中间物质用来传 递DNA上的信息。他们研究大肠杆菌中与乳糖代谢有关酶类的生物合成时发现诱导物,如 异丙基硫代半乳糖苷(B-isopropyIthiogalactoside)的加入,可以立刻使酶蛋白的合成速度增 加上千倍。而诱导物一旦消失,又可使酶蛋白的合成立刻停止。这个实验结果给人的启示 是:蛋白质合成的模板是一种不稳定的物质,其半寿期很短。他们对这种信使物质的性质 作了如下的预言: ①信使是一种多核苷酸。 ②信使的碱基组成应与相应的DNA的碱基组成相一致。 298
298 酵母线粒体、原生动物纤毛虫也有类似情形。所以结论应该是:遗传密码并非是绝对 通用的,而是近于完全通用的。 第二节 蛋白质合成的分子基础 氨基酸是在核糖体上加入到多肽链中的。在与mRNA作用之前,氨基酸先共价地与转 运RNA(transfer RNA,tRNA)形成氨酰-tRNA。氨酰-tRNA结合到mRNA的特殊位点上。 mRNA含有遗传密码的信息,用于指导特定氨基酸序列多肽链的合成。一个核糖体结合到 一个mRNA分子合成起始序列上,并由此开始读码,沿着密码序列合成一条多肽链。读码 的方向是从mRNA的5′到3′,而合成出来的多肽则是从氨基端到羧基端。通常,一个 mRNA分子上可结合有多个不同时间开始翻译的核糖体,这样的结构称为多聚核糖体 (polysome)。在原核细胞中,mRNA的转录与多肽的翻译是同时进行的,如图12-1所示,可 见在染色体DNA分子上,有多条正在转录的mRNA分子,每个分子上都结合有多个正在进 行翻译的核糖体。真核生物的转录与翻译是在不同的地方进行的,核糖体可以自由地存在 于细胞质中,或者与内质网膜结合。RNA分子在蛋白质的合成中起着举足轻重的作用,除 了mRNA和tRNA,核糖体还含有核糖体RNA(ribosome RNA, rRNA)。 一、mRNA是蛋白质合成的模板 Jacob F.和Monod J.早在1961年就已提出mRNA的概念。他们认为,既然蛋白质是在细胞质 中合成的,而编码蛋白质的信息载体DNA却在细胞核内,所以必定有一种中间物质用来传 递DNA上的信息。他们研究大肠杆菌中与乳糖代谢有关酶类的生物合成时发现诱导物,如 异丙基硫代半乳糖苷(β-isopropylthiogalactoside)的加入,可以立刻使酶蛋白的合成速度增 加上千倍。而诱导物一旦消失,又可使酶蛋白的合成立刻停止。这个实验结果给人的启示 是:蛋白质合成的模板是一种不稳定的物质,其半寿期很短。他们对这种信使物质的性质 作了如下的预言: ①信使是一种多核苷酸。 ②信使的碱基组成应与相应的DNA的碱基组成相一致。 密码子 通常情况下可编码的 线粒体DNA所编码的 UGA UGG AUA AUG AGA AGG 终止信号 色氨酸 异亮氨酸 甲硫氨酸 精氨酸 精氨酸 色氨酸 色氨酸 甲硫氨酸 甲硫氨酸 终止信号 终止信号 表 12-6 人线粒体 DNA 中密码编制特点

③信使的长度应是不同的,因为由它们所编码的多肽链的长度是不同的。 ④在多肽合成时信使应与核糖体作短暂的结合。 连有多个核籍体的mRNA 翻详的方向 8 06 c2 DNA 转录的方向 图12-1大肠杆菌中mRNA的转录与多肽的翻译是同时进行的 A为多聚核糖体的电镜照片 ⑤信使的半寿期很短,所以信使的合成速度应该是很快的。 所以,这样的信使可能是一种RNA。但是当时己发现的两种RNA都不具备这些特性。 各种生物的RNA的大小差异不大,碱基组成的变化也不大。tRNA除了有与rRNA相同的问 题以外,它们的分子也太小,所以这两种RNA都不能胜任信使的功能。可喜的是当时己有 人提出过,认为细胞中有可能存在第三种RNA。被噬菌体T感染后的大肠杆菌中,有人发 现有一种新的RNA,它的代谢速度极快,分子的大小也参差不齐,碱基组成又与TDNA相 致。这些特性都符合信使分子的要求 mRNA的概念提出后,还必须要用实验来证明这种概念是否正确。Brenner S.,Jacob F. 和Monod M.等人设计了一组实验:用噬菌体T感染大肠杆菌后,发现几乎所有在细胞内合 成的蛋白质都不再是细胞本身的蛋白质,而是噬茵体所编码的蛋白质,这些蛋白质的合成 速度与细胞总RNA的合成速度无关:T感染后不久,细胞中出现了少量半寿期很短的RNA 它们的碱基组成与TDNA是一致的。上述这些特性都与他们预言的信使分子特性十分符 合。他们将大肠杆菌接种在含有重标记(5N和PC)的培养基上,再用T感染。感染后立刻将 29g
299 ③信使的长度应是不同的,因为由它们所编码的多肽链的长度是不同的。 ④在多肽合成时信使应与核糖体作短暂的结合。 图12-1 大肠杆菌中mRNA的转录与多肽的翻译是同时进行的 A为多聚核糖体的电镜照片 ⑤信使的半寿期很短,所以信使的合成速度应该是很快的。 所以,这样的信使可能是一种RNA。但是当时已发现的两种RNA都不具备这些特性。 各种生物的rRNA的大小差异不大,碱基组成的变化也不大。tRNA除了有与rRNA相同的问 题以外,它们的分子也太小,所以这两种RNA都不能胜任信使的功能。可喜的是当时已有 人提出过,认为细胞中有可能存在第三种RNA。被噬菌体T2感染后的大肠杆菌中,有人发 现有一种新的RNA,它的代谢速度极快,分子的大小也参差不齐,碱基组成又与T2DNA相 一致。这些特性都符合信使分子的要求。 mRNA的概念提出后,还必须要用实验来证明这种概念是否正确。Brenner S.,Jacob F. 和Monod M.等人设计了一组实验:用噬菌体T2感染大肠杆菌后,发现几乎所有在细胞内合 成的蛋白质都不再是细胞本身的蛋白质,而是噬菌体所编码的蛋白质,这些蛋白质的合成 速度与细胞总RNA的合成速度无关;T2感染后不久,细胞中出现了少量半寿期很短的RNA, 它们的碱基组成与T2DNA是一致的。上述这些特性都与他们预言的信使分子特性十分符 合。他们将大肠杆菌接种在含有重标记( 15N和12C)的培养基上,再用T2感染。感染后立刻将
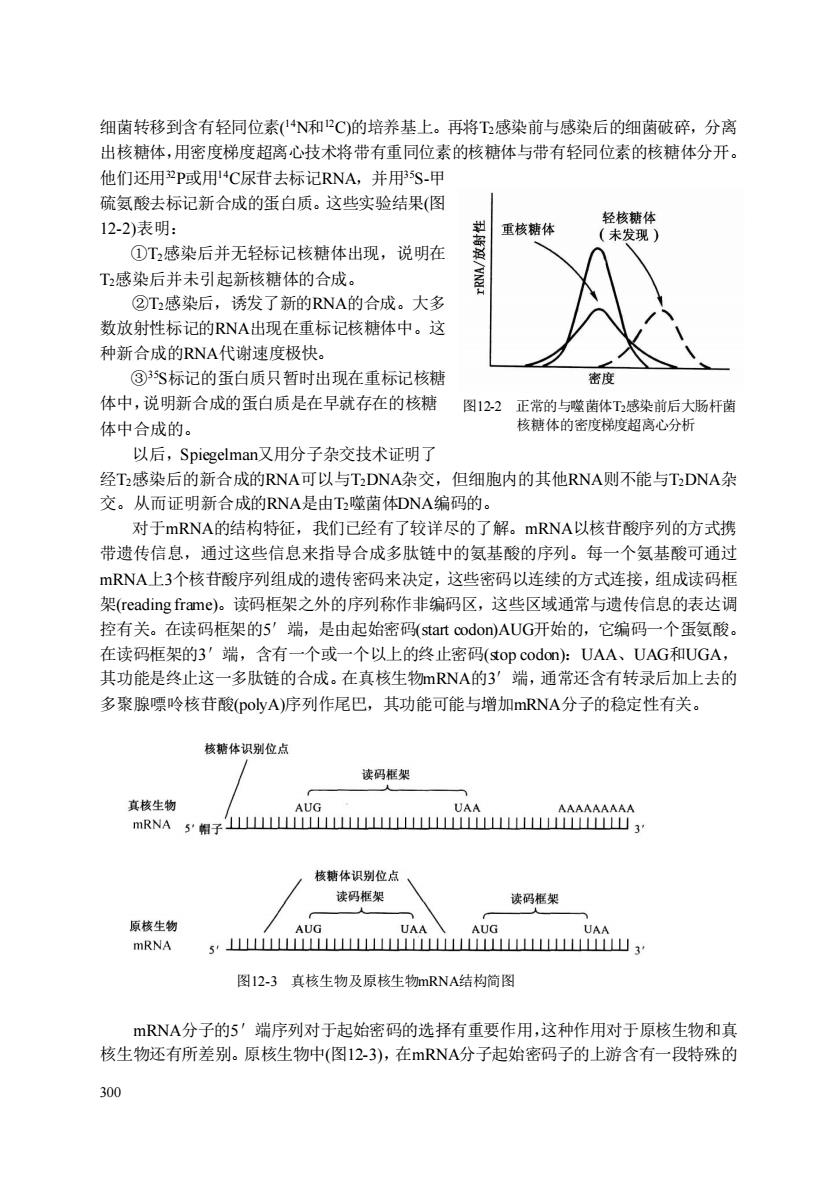
细菌转移到含有轻同位素(N和P℃)的培养基上。再将T感染前与感染后的细菌破碎,分离 出核糖体,用密度梯度超离心技术将带有重同位素的核糖体与带有轻同位素的核糖体分开。 他们还用2P或用4C尿苷去标记RNA,并用5S-甲 硫氨酸去标记新合成的蛋白质。这些实验结果(图 12-2)表明: 重核糖体 ①T2感染后并无轻标记核糖体出现,说明在 T2感染后并未引起新核糖体的合成。 ②T感染后,诱发了新的RNA的合成。大多 数放射性标记的RNA出现在重标记核糖体中。这 种新合成的RNA代谢速度极快。 ③S标记的蛋白质只暂时出现在重标记核糖 密度 体中,说明新合成的蛋白质是在早就存在的核糖图12.2正常的与噬菌体T感染前后大肠杆菌 体中合成的。 核糖体的密度梯度超离心分析 以后,Spiegelman又用分子杂交技术证明了 经T感染后的新合成的RNA可以与TDNA杂交,但细胞内的其他RNA则不能与TDNA杂 交。从而证明新合成的RNA是由T噬菌体DNA编码的。 对于mRNA的结构特征,我们已经有了较详尽的了解。mRNA以核苷酸序列的方式携 带遗传信息,通过这些信息来指导合成多肽链中的氨基酸的序列。每一个氨基酸可通过 mRNA上3个核苷酸序列组成的遗传密码来决定,这些密码以连续的方式连接,组成读码框 架(reading frame)。读码框架之外的序列称作非编码区,这些区域通常与遗传信息的表达调 控有关。在读码框架的5'端,是由起始密码((start codon)AUG开始的,它编码一个蛋氨酸 在读码框架的3'端,含有一个或一个以上的终止密码(top codon:UAA、UAG和UGA, 其功能是终止这一多肽链的合成。在真核生物mRNA的3'端,通常还含有转录后加上去的 多聚腺票岭核苷酸(Do小vA)序列作尾巴,其功能可能与增加mRNA分子的隐定性有关。 核糖体识别位店 读码框架 真核生物 mRNA 5帽子山YMM 核糖体识别位点 读码框架 读码据架 原核生物 mRNA s'出i山, 图12-3真核生物及原核生物mRNA结构简图 mRNA分子的5'端序列对于起始密码的选择有重要作用,这种作用对于原核生物和真 核生物还有所差别。原核生物中(图12-3),在mRNA分子起始密码子的上游含有一段特殊的 300
300 细菌转移到含有轻同位素( 14N和12C)的培养基上。再将T2感染前与感染后的细菌破碎,分离 出核糖体,用密度梯度超离心技术将带有重同位素的核糖体与带有轻同位素的核糖体分开。 他们还用32P或用14C尿苷去标记RNA,并用35S-甲 硫氨酸去标记新合成的蛋白质。这些实验结果(图 12-2)表明: ①T2感染后并无轻标记核糖体出现,说明在 T2感染后并未引起新核糖体的合成。 ②T2感染后,诱发了新的RNA的合成。大多 数放射性标记的RNA出现在重标记核糖体中。这 种新合成的RNA代谢速度极快。 ③35S标记的蛋白质只暂时出现在重标记核糖 体中,说明新合成的蛋白质是在早就存在的核糖 体中合成的。 以后,Spiegelman又用分子杂交技术证明了 经T2感染后的新合成的RNA可以与T2DNA杂交,但细胞内的其他RNA则不能与T2DNA杂 交。从而证明新合成的RNA是由T2噬菌体DNA编码的。 对于mRNA的结构特征,我们已经有了较详尽的了解。mRNA以核苷酸序列的方式携 带遗传信息,通过这些信息来指导合成多肽链中的氨基酸的序列。每一个氨基酸可通过 mRNA上3个核苷酸序列组成的遗传密码来决定,这些密码以连续的方式连接,组成读码框 架(reading frame)。读码框架之外的序列称作非编码区,这些区域通常与遗传信息的表达调 控有关。在读码框架的5′端,是由起始密码(start codon)AUG开始的,它编码一个蛋氨酸。 在读码框架的3′端,含有一个或一个以上的终止密码(stop codon):UAA、UAG和UGA, 其功能是终止这一多肽链的合成。在真核生物mRNA的3′端,通常还含有转录后加上去的 多聚腺嘌呤核苷酸(polyA)序列作尾巴,其功能可能与增加mRNA分子的稳定性有关。 图12-3 真核生物及原核生物mRNA结构简图 mRNA分子的5′端序列对于起始密码的选择有重要作用,这种作用对于原核生物和真 核生物还有所差别。原核生物中(图12-3),在mRNA分子起始密码子的上游含有一段特殊的 图12-2 正常的与噬菌体T2感染前后大肠杆菌 核糖体的密度梯度超离心分析

核糖体结合位点(ribosome-binding site序列,这一结合位点使得核糖体能铭识别正确的起始 密码AUG。原核生物的mRNA通常是多基因的,分子内的核糖体结合位点使得多个基因可 独立地进行读码框架的翻译,得到不同的蛋白质。而对于真核生物而言,其mRNA通常只 为一条多肽链编码,核糖体与mRNA5'端的核糖体进入部位((ibosome enry site)结合之后 通过一种扫描机制向3'端移动来寻找起始密码,mRNA5'末端的帽子结构可能对于核糖 体进入部位的识别起着一定作用。翻译的起始通常开始于从核糖体进入部位向下游扫描到 的第一个AUG序列。 二、tRNA转运活化的氨基酸至mRNA模板上 tRNA含有两个关键的部位:一个是氨基酸结合部位,另一个是与mRNA的结合部位。 对于组成蛋白质的20种氨基酸来说,每一种至少有一种RNA来负责转运。为了准确地翻译 每一种RNA必需能被很好地识别。在书写时,将所运氨基酸写在tRNA的右上角,如RNAc 及tRNA如分别表示苯丙氨酸和丝氨酸转运的tRNA。大多数氨基酸具有几种用来转运的 tRNA,一个细胞中,通常含有50或更多的不同的 RNA分子 tRNA是第一个通过X射线品体衍射技术测定了RNA分子的空间结构,其他tRNA都 与它类似,可用一个如图12-4的结构来表征其一级、二级、三级结构。所有的RNA都是有 由5095个核苷酸组成的一条多聚核苷酸链,这条链经过折叠,呈现三叶草型结构,含有4 个双链的茎和4个单链的环。5'端和3'端的碱基通过形成7个Waston-Crick碱基配对将两 端拉到一起,形成受体端,氨基酸通过与3'端的核糖连接而形成氨酰RNA分子。tRNA 的3'端通常是CCA的序列。 OH
301 核糖体结合位点(ribosome-binding site)序列,这一结合位点使得核糖体能够识别正确的起始 密码AUG。原核生物的mRNA通常是多基因的,分子内的核糖体结合位点使得多个基因可 独立地进行读码框架的翻译,得到不同的蛋白质。而对于真核生物而言,其mRNA通常只 为一条多肽链编码,核糖体与mRNA 5′端的核糖体进入部位(ribosome entry site)结合之后, 通过一种扫描机制向3′端移动来寻找起始密码,mRNA 5′末端的帽子结构可能对于核糖 体进入部位的识别起着一定作用。翻译的起始通常开始于从核糖体进入部位向下游扫描到 的第一个AUG序列。 二、tRNA转运活化的氨基酸至mRNA模板上 tRNA含有两个关键的部位:一个是氨基酸结合部位,另一个是与mRNA的结合部位。 对于组成蛋白质的20种氨基酸来说,每一种至少有一种tRNA来负责转运。为了准确地翻译, 每一种tRNA必需能被很好地识别。在书写时,将所运氨基酸写在tRNA的右上角,如tRNAPhe 及tRNASer分别表示苯丙氨酸和丝氨酸转运的tRNA。大多数氨基酸具有几种用来转运的 tRNA,一个细胞中,通常含有50或更多的不同的 tRNA分子。 tRNAPhe是第一个通过X射线晶体衍射技术测定了tRNA分子的空间结构,其他tRNA都 与它类似,可用一个如图12-4的结构来表征其一级、二级、三级结构。所有的tRNA都是有 由50-95个核苷酸组成的一条多聚核苷酸链,这条链经过折叠,呈现三叶草型结构,含有4 个双链的茎和4个单链的环。5′端和3′端的碱基通过形成7个Waston-Crick碱基配对将两 端拉到一起,形成受体端,氨基酸通过与3′端的核糖连接而形成氨酰-tRNA分子。tRNA 的3′端通常是CCA的序列

图124RNA的结构简图 未配对的环的命名由其特定的结构来定。环的大小在 7~11个核苷酸之间,常含有稀有碱基dihydrouracil,故命名 为D环:环Ⅱ含有被称作反密码子(anticodon)的个碱基序列, 被命名为反密码子环。RNA的这一部分在蛋白质的合成中非 常重要,它可与mRNA模板上的密码子进行碱基配对的专 性的识别,并将所携带的氨基酸送入到合成的多肽链的指定 位置上:环Ⅲ是可变环,其组成可在321个碱基之间,是 C O tRNA大小变化最大的区域:环V含有稀有的胸腺嘧啶核糖核 mRNA 苷(ribothymidine)和假尿嘧啶核苷(pseudouridine)(符号中表 示)碱基作为不变序列,故把它叫做T中C环。 图12-5斋码子与反密码子 tRNA三叶草型的二级结构可折叠成L型的三维结构,如图12-4所示,这一结构由两个 螺旋以直角的方位构成,结合氨基酸的一端称接受臂(accepor amm),另一端则含有反密码 子,被称作反密码子臂(anticodon arm。tRNA分子上与多肽合成有关的位点至少有4个,分 别为3'端CCA上的氨基酸接受位点、识别氨酰RNA合成酶的位点、核糖体识别位点及反 密码子位点。 RNA在识别mRNA分子上的密码子时,具有接头adaptor)的作用。氢基酸一旦与RNA 形成氨酰tRNA后,进一步的去向就由tRNA来决定了。tRNA凭借自身的反密码子与mRNA 分子上的密码子相识别图12-5),而把所带的氨基酸送到肽链的一定位置上。Chapeville及 Lipmann(1962)做了一个巧妙的实验来证明这一点。将放射性同位素标记的半胱氨酸在半胱 氨酰-RNA合成酶催化下与RNA形城半胱氨酰-tRNA,然后用活性镍作催化剂,使半 胱氨酸转变成丙氨酸,形成丙氨酰RNA9:。然后将它放到网织红细胞无细胞体系中进行 蛋白质合成。分析后,发现丙氨酸插入了本应由半胱氨酸所占的位置 前面在讨论遗传密码的性质时曾提到过密码的简并性问题,这里将进一步讨论与此有关 的tRNA分子突变与校正基因(suppressor gene)的问题。遗传学家早就发现了回复突变 (reverse mutation)现象。回复突变的原因很多,其中有一种回复突变是由于其在基因上发生 的 一个突变引起的,这称为基因间校正突变。长期以来人们很难解释基因间校正突变。但 是现在由于对RNA的结构功能有了较深入的了解,基因间校正突变的本质己经被揭露了。 大多数校正突变是发生在RNA基因的突变上,从而使tRNA的反密码子在阅读mRNA的信 息时发生了变化。下面举一个例子加以说明(图12-6)。 H,'N- 有活力的多肽 GAG(Gln)到 UAG终止)的突出 HN C00 第变使鉴出读作Tr 无活力的多肽 有活力的突变体多肽 图12-6基因间校正突变图解 302
302 图12-4 tRNA的结构简图 未配对的环的命名由其特定的结构来定。环I的大小在 7~11个核苷酸之间,常含有稀有碱基dihydrouracil,故命名 为D环;环Ⅱ含有被称作反密码子(anticodon)的3个碱基序列, 被命名为反密码子环。tRNA的这一部分在蛋白质的合成中非 常重要,它可与mRNA模板上的密码子进行碱基配对的专一 性的识别,并将所携带的氨基酸送入到合成的多肽链的指定 位置上;环Ⅲ是可变环,其组成可在3~21个碱基之间,是 tRNA大小变化最大的区域;环Ⅳ含有稀有的胸腺嘧啶核糖核 苷(ribothymidine)和假尿嘧啶核苷(pseudouridine)(符号ψ表 示)碱基作为不变序列,故把它叫做TψC环。 tRNA三叶草型的二级结构可折叠成L-型的三维结构,如图12-4所示,这一结构由两个 螺旋以直角的方位构成,结合氨基酸的一端称接受臂(acceptor arm),另一端则含有反密码 子,被称作反密码子臂(anticodon arm)。tRNA分子上与多肽合成有关的位点至少有4个,分 别为3′端CCA上的氨基酸接受位点、识别氨酰-tRNA合成酶的位点、核糖体识别位点及反 密码子位点。 tRNA在识别mRNA分子上的密码子时,具有接头(adaptor)的作用。氨基酸一旦与tRNA 形成氨酰-tRNA后,进一步的去向就由tRNA来决定了。tRNA凭借自身的反密码子与mRNA 分子上的密码子相识别(图12-5),而把所带的氨基酸送到肽链的一定位置上。Chapeville及 Lipmann(1962)做了一个巧妙的实验来证明这一点。将放射性同位素标记的半胱氨酸在半胱 氨酰-tRNA合成酶催化下与tRNACys形成半胱氨酰-tRNACys,然后用活性镍作催化剂,使半 胱氨酸转变成丙氨酸,形成丙氨酰-tRNACys。然后将它放到网织红细胞无细胞体系中进行 蛋白质合成。分析后,发现丙氨酸插入了本应由半胱氨酸所占的位置。 前面在讨论遗传密码的性质时曾提到过密码的简并性问题。这里将进一步讨论与此有关 的tRNA分子突变与校正基因(suppressor gene)的问题。遗传学家早就发现了回复突变 (reverse mutation)现象。回复突变的原因很多,其中有一种回复突变是由于其在基因上发生 的一个突变引起的,这称为基因间校正突变。长期以来人们很难解释基因间校正突变。但 是现在由于对tRNA的结构功能有了较深入的了解,基因间校正突变的本质已经被揭露了。 大多数校正突变是发生在tRNA基因的突变上,从而使tRNA的反密码子在阅读mRNA的信 息时发生了变化。下面举一个例子加以说明(图12-6)。 图 12-5 密码子与反密码子 的识别 H3 +N————————Gln————————COO- 有活力的多肽 GAG(Gln)到 UAG(终止)的突出 H3 +N————————COO- 第二个突变使 tRNATyr 无活力的多肽 可以将 UAG(终止)读作 Tyr H3 +N————————Tyr————————COO- 有活力的突变体多肽 图 12-6 基 因间校正突变图解