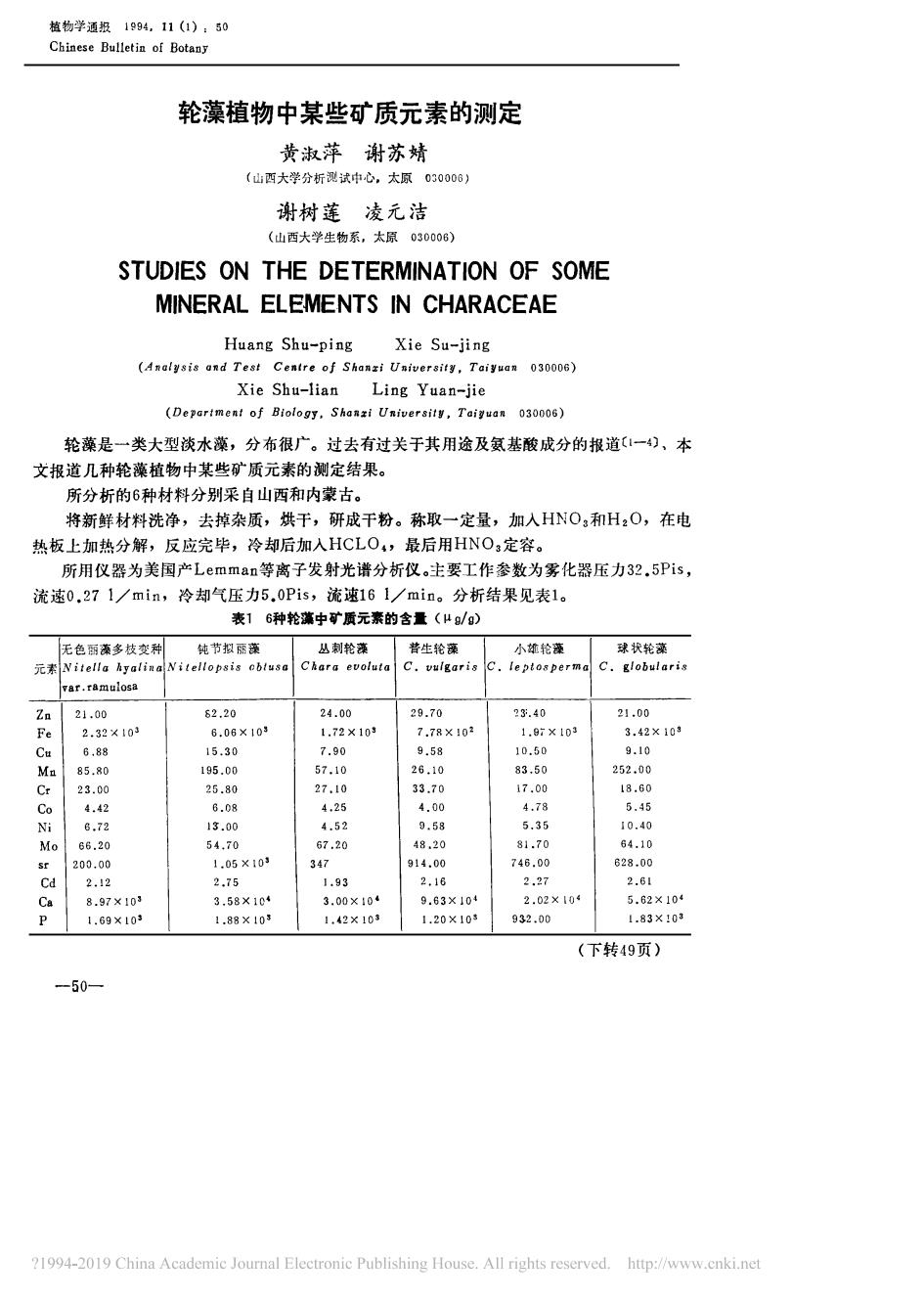
植物学通报1994,11(1):50Chinese Bulletin of Botany轮藻植物中某些矿质元素的测定黄淑萍谢苏婧(山西大学分析测试中心,太原030006)谢树莲凌元洁(山西大学生物系,太原030006)STUDIESONTHEDETERMINATIONOFSOMEMINERALELEMENTSINCHARACEAEHuang Shu-pingXie Su-jing(Analgsis andTest Cenire of Shanzi University,Taiyuan 030006)Xie Shu-lianLing Yuan-jie(Department of Biology,Shanzi University,Taiguan 030006)轮藻是一类大型淡水藻,分布很广。过去有过关于其用途及氨基酸成分的报道(1一4)、本文报道几种轮藻植物中某些矿质元素的测定结果。所分析的6种材料分别采自山西和内蒙古。将新鲜材料洗净,去掉杂质,烘干,研成干粉。称取一定量,加入HNO,和H2O,在电热板上加热分解,反应完毕,冷却后加入HCLO4,最后用HNO:定容。所用仪器为美国产Lemman等离子发射光谱分析仪。主要工作参数为雾化器压力32.5Pis,流速0.271/min,冷却气压力5.0Pis,流速161/min。分析结果见表1。表16种轮藻中矿质元素的含量(μg/g)无色丽藻多枝变种钝节拟丽藻丛刺轮藻普生轮藻小雄轮藻球状轮藻元素 Nitella hyalina Nitellopsis obtusa Chara evolutaC.globularisC.vulgarisc.leptospermalvar.ramulosa29.7023.4021.00Zn21.0062.2024.006.06X1033.42×10gFe2.32X1031.72×10g7.78X1021.97X1039.5810.509.10Cu6.8815.307.9026.1083.50252.00Mn85,80195.0057.1027.1033.7017.0018.60Cr23.0025.804,254.004.785.45Co4.426.084.526.7213.009.585.3510.40Ni54.7067.2048.2081.7064.10Mo66.201.05×103347914.00746.00628.00200.00sr1.932,162.272.61Cd2.122.75Ca3.58×1043.00×10*9.63X1042.02×1045.62×1048.97×1031.42X108932.001.83X103P1.69×1031.88X1031.20 ×10s(下转49页)-50—21994-2019China Academic Journal Electronic PublishingHouse.Allrights reserved.http://www.cnki.net
植物学通 报 1 9 9 4 , 1 2 ( l ) : 5 0 C h还 e s e B u l l e ti n o f B o t a n y 轮藻植物中某些矿质元素的测定 黄淑萍 谢苏蜻 ( 山西 大学分析测 试中心 , 太原 。 3 0 0 0 勺 谢树 莲 凌 元 洁 ( 山西大学生物系 , 太原 。 3。 0 6 ) S T UD IES O N T HE D E T E RM!N AT 10 N OF SO ME M!N ER AL E LE ME NT S IN C HAR AC E AE H u a n g S h u 一 P i n g X i e S u 一 j i n g ( A , a l 夕s 艺。 a , d T e s t C e . l r e o f S h a : 二 1 U o f o e r 。 ` t犷 , T a ` 李“ a : 0 3 0 0 0 6 ) X i e S h u 一 l i a n L i n g Y u a n 一j i e ( D e , 。 r t仇 e o t o f B i o l o 夕 y , S h a 泥 劣 i U 二 `p e r s i t梦 . T a `梦。 a : 0 3 0 0 0 6 ) 轮藻是一类大型 淡水 藻 , 分布很广 。 过去 有过关 于其用途及 氨基酸 成分 的 报道 〔`一4 〕 、 本 文报道 几种 轮藻植物 中某些矿质元 素的 测定结果 。 所分 析的 6种材料 分别采 自山西和 内蒙古 。 将新鲜材料洗 净 , 去 掉杂 质 , 烘干 , 研成干 粉 。 称取一 定量 , 加 入 H N O 3和 H Z O , 在电 热板 上加 热 分解 , 反 应完 毕 , 冷却后加入 H C L O ; , 最后 用 H N 0 3 定容 。 所 用仪 器为 美国产 L e m m a n 等离子发 射光谱 分析仪 。 主 要工作 参数 为雾 化器压 力 32 . S iP s , 流 速 0 . 2 7 1 / m i n , 冷却 气压 力 5 。 OP i s , 流速 1 6 1/ m i n 。 分 析结果 见表 1 。 表 1 6种轮燕 中矿质元紊的 含l ( “ 。 / 。 ) 钝节拟 丽 藻 元素 无 色丽 藻多枝 变种 N £t e l l a h y a l£n a v a r . r a m u l o s a N 艺t e l l o P s 艺5 o b t u s a 丛刺轮藻 C h a r a e 口 o l u t a 普生轮 藻 C . t, u l g a r 艺s 小雄轮藻 C . I e 夕亡o s P e r 仍 a 球状轮藻 C . g l o b “ l a r 蕊s 至 . 0 0 . 7 2 X 1 0 3 . 9 0 。 1 0 . 1 0 . 2 5 . 5 2 . 2 0 2 1 . 0 0 5 2 . 2 0 2 4 . 0 0 2 9 。 7 0 2 :3 . 4 0 2 . 3 2 义 1 0 3 6 . 0 6 x l 0 3 1 . 7 2 X 1 0 3 7 . 7 8 X 1 0 2 1 . 9 7 X 1 0 3 6 . 8 8 1 5 . 3 0 7 . 9 0 9 。 5 8 1 0 . 5 0 8 5 . 8 0 1 9 5 . 0 0 5 7 。 1 0 2 6 。 1 0 8 3 . 5 0 2 3 . 0 0 2 5 . 8 0 2 7 . 1 0 3 3 . 7 0 1 7 . 0 0 4 。 4 2 6 . 0 8 4 . 2 5 4 。 0 0 4 . 7 8 6 。 7 2 l了 。 0 0 4 . 5 2 9 . 5 8 5 . 3 5 6 6 . 2 0 5 4 . 7 0 6 7 . 2 0 4 8 . 2 0 8 1 。 7 0 2 0 0 . 0 0 1 . 0 5 X 1 0 3 3 4了 9 1 4 。 0 0 7 4 6 . 0 0 2 。 1 2 2 。 7 5 1 . 9 3 2 . 1 6 2 。 2 7 8 . 9 7 X 1 0 3 3 . 5 8 义 1 0 4 3 . 0 0 x 1 0 . 9 . 6 3 X 1 0 4 2 . 0 2 又 1 0 4 1 . 6 9 X I O , 1 . 8 8 x 1 0 , 1 . 4 2 义 1 0 5 1 . 2 0 X 1 0 s 9 3 2 . 0 0 ( 下 转 4 9页 ) 一 5 0一

表1不同光质处理下稻苗经高温伤害和恢复过程中SOD活性(酶单位·g)与MDA含量(μM9W)的变化DRB1GW(ck)SODMDASODMDASODMDASODMDASODMDASODMDA不同光质下正常生长145.508.45167.308.26138.209.48819.107.16218.206.13181.807,74高温处理第一天94.1710.67115.539.8080.2911.27130.158.98182.157.56123.699.47第二天100.8910.6972.4712.0265.3912.05133.819.69174.208.65 115.8810.15温室下恢第一天50.8513.6294.2612.3650.9112.65123.059.70181.5610.497.99108.76115.8310.9272.7312.13129.84复生长第二天72.68 12.069.28189.097.36123.0510.21第三天101.5311.45138.3210.2086,8211.49147,348.89219.705.97130.889.35的修复有一个迟滞期。直至解除逆境后的第二天,各处理幼苗SOD活性才有不同程度的上升,MDA含量才下降。各处理幼苗SOD活性的降升与MDA含量的升降呈显著负相关,相关系数分别为:rp=-0.9710**,rn=-0.8789*,ry=-0.9836**,ra=-0.9784**,rg=-0.9828一经LSR测验,二者显著相关,。。“**,rw=-0.9674**(","一经LSIZ测验,二者极显著相关。),从而进一步证实高温对水稻幼苗的伤害是由于高温抑制了SOD活性,破坏了水稻幼苗体内自由基的产生与清除的动态平衡关系,加速了膜脂的过氧化作用,导致膜结构和功能的破坏,影响植物的正常生长。因此可以认为,SOD在植物抵抗高温伤害的过程中,可能起着防止、中断和终止膜脂过氧化对细胞膜系统损伤的酶性保护作用(1)。从表中的结果还可以看出:(1)原培养于不同光质下的水稻幼苗耐高温能力有较大差异。其中以原蓝光处理的稻苗抗高温能力最强,黄光最弱。(2)室温下恢复生长到第三天,只有原蓝光下培育的稻苗SOD活性、MDA含量恢复到原来(未经高温处理前)水平,幼苗生长已正常,其它光质处理的稻苗均还未恢复到正常生长,说明原蓝光处理的稻苗恢复能力也最强。产生以上差异的原因可能与不同光质下所形成的酶性保护系统的强弱、产生自由基能力的大小以及培养期间所形成的“物质基础”不同有关,详细机理有待进一步探讨。参考文献【1)方允中,牵书杰主编,1989,自由基与基础理论及其在生物学和层学中的应印,科学出吸社至41一45可【2)邵从本等、1983,植物生理学通证,(5):4e49【3】林植芳等,1984,植物学报,26(6)3605—615,[4]Coffman,F.A.,1957,AgronomyJ.,49368-373.[5]Rabinowitch,H.D.andI.Fridovici,1985,Planta,164:524--528(上接第50页)参考文献[1]丁恒山,1987,中国的药用孢子植物,科学技术出版社,上海。[2】谢树莲、凌元活,1991,山西大学学报(自然科学版),14(1),88—92[3] Imahori, K.,1954,Ecclogy,phytogeography and taxonomy of the Japanese Charophyta,kanazawaUniversity,Japan[4]ZaneveldJ.S.,1940,Bluma,4(1):1-224.-49-21994-2019 China Academic Journal Electronic Publishing House.All rights reserved,http://www.cenki.net
表 1 不 同光质 处理下 稻苗经高温 伤害 和恢复过程中 sD0 活性 (酶单位 · 。硫 )与M AD 含 t ( , .M g豁 ) 的 变化 D R G B W ( e k ) S O D M D A M D A M D A M D A S O D M D A S O D M D人 不 同 高温 1 4 5 . 5 0 8 . 4 5 9 4 . 1 7 1 0 . 6 7 7 夕 . 4 7 ] 2 . 0 2 弓0 . 8 5 1 3 . 6 2 7 2 。 6 8 1 2 . 0 6 1 0 ] . 5 3 1 1 。 4 5 S O D 1 G7 . 3 O 1 1 5 . 5 3 1 0 0 . 8 9 9 4 . 2 6 1 1 5 . 8 3 1 3 8 . 3 2 SO D 1 3 8 . 2 0 3 0 . 2 9 6 5 . 3 9 5 0 . 9 1 7 2 . 7 3 8 6 。 8 2 S O D 9 . 1 0 7 . 1 6 1 3 0 . 1 5 8 . 9 8 1 2 3 . 8 ! 9 . 6 9 1 2 3 . 0 5 9 . 7 0 1? 9 . 8 4 9 . 2 8 1 4 7 . 3 4 8 . 8 9 2 1 8 . 2 0 6 . 1 3 1 8 1 . 8 0 1 8 2 . 1 5 7 . 5 6 1 2 3 . 6 9 1 7 4 . 2 0 8 . 6 5 1 1 5 . 8 8 1 8 1 . 5 6 7 . 9 9 1 0 8 . 7 6 [ 8 9 . 0 9 7 . 3 G 1 2 3 . 0 5 夕1 9 . 7 0 5 . 9 7 1 3 0 . 8 8 : :: 79 温室下恢 复生 长 第 一天 第 二天 第 三天 1 0 . 1 5 1 0 。 4 9 1 0 。 2 1 9 . 3 5 叮48 .05 “ :13 比.912 .12 扰80 12二 .9G.36 120.108.9 的 修复有 一个 迟滞期 。 直 至解除逆境后 的第二夭 , 各 处理 幼 苗 S O D 活性 才 有不同程 度的上 升 , M D A 含量 才 下降 。 各处理 幼苗 S O D 活性 的 降升与M D A 含量 的升 降 呈显 著负相 关 , 相 关 系数 分 别为 : r 。 = 一 0 . 9 7 1 0令 . , r R = 一 0 . 8 7 8 9 . , r y = 一 0 . 9 8 3 6二 , 1 · 。 = 一 0 . 9 7 5 4 , 气 r 。 = 一 0 . 9 5 2 5 . 气 r w = 一 0 . 9 6 7 4二 ( “ ` ’ 一 经 SL R测 甄 二 者显著相关 : ` 二 ” 一 里儿 slz 测 验 · 二 者极显 著相关 . ) , 从而进 一步 证实高温 对 水稻 幼 苗的 伤 害是 由于高温 抑制 了 S O D 活 性 , 破 坏 了 水 稻 幼 苗 体 内 自由基 的产 生 与清除的 动态 乎衡 关系 , 加 速 了膜 脂 的过 氧化作 用 , 导致 膜 结构 和 功 能的破 坏 , 影响 植物的 正常 生 长 。 因此 可以认 为 , S O D 在植物 抵抗 高温 伤 害的 过 程 中 , 可能起 着 防 i L 、 中断 和终 止膜 脂过 氧化对 细 胞膜 系统损 伤的 酶性 保护 作 用〔 ,〕 。 从表 中的结果还 可以 看 出 : ( 1) 原 培 养于不 同 光质 下的 水稻 幼苗 耐 高温 能 力有 较 大 差 异 。 其 中以原 蓝光处 理 的稻 苗抗 高温能 力最强 , 黄光最弱 。 ( 2) 室温 下恢复生 长到 第三 天 , 只有原 蓝 光下 培 育的稻 苗 S O D 活性 、 M D A 含量 恢 复到 原 来 ( 未经 高温处理 前 ) 水平 , 幼 苗 生 长 已正 常 , 其它光 质处 趣的 稻苗 均还 未恢复 到正常 生 长 , 说明原 蓝光处 理 的稻 苗 恢复能 力 也 最 强 。 产生 以上 差异 的原 因可能 与不 同光 质下所 形成的 酶性保护系统 的 强弱 、 产 生 自由基 能 力 的 大小 以 及培 养期间 所形 成的 “ 物 质基 础 ” 不 同有关 , 详细机 理 有待进一 步探 讨 。 参 考 文 献 〔 l 〕 方允 巾 、 李书杰 主编 , 1 9 8 9 , 自由基 与酶 墓 触 理论 及 其 在生 物 学和 医学巾的 应 用 一 科 学出 版 社 节 魂! 一朽 而 ( 2 〕 邵从本等 、 王9韶 , 植物生理 学通 讯 , ( 5 ) : 4 e 一49 〔 3 〕 林 植芳等 1 9已4 , 植物学报 , 2 6 ( G ) , 6 0 5一 6 一5 . 〔 4 〕 C o f f m a n , F . A . , 1 9 5 7 , A g r o n o m y J . , 4 9 : 3 6 8一 3 7 3 . 〔 5 」 R a b i n o w i t e 卜 , H . D . a n d I . F r i d o v i 。 与 , 一9 3 弓 , P l a n t a , x . 4 : 5 2 4一5 2 8 . ( 上接第50 页 ) 参 考 文 献 〔l] 丁 恒 山 , 1 9 87 , 中国 的 药用 抱子 植物 , 科学技术出版社 , 上海 . [ 2 〕 谢 树莲 、 凌元 洁 , l 。 l , 山 西大学学报 ( 自然科学版) , 24 ( 一) : 日s一。 2 . 〔, ] Im a b o r i K . , 1 9 5 4 E e o l o g y , p h y t o g e o g r a p il 了 a n d t a x o n o m y o f t卜e J a Pa n e s e C h a r o p h y t a , 卜a o a Z a w a U n i v e r s i t y , J a P a n . [ 4 〕 Z a n e v e l d J . 5 . , 1 0 4 0 , B l u 饥 a , 4 ( 1 ) : 1一 2 2 4 。 一4 9一