
第一章啤酒,面包,奶酪■ 假丝时母(Candida 拟内跑霉(Endomycopsis) 单细胞酵母(Saccharomycs 青霉素 (Penicillium) 球拟酵母属(Torulopsis) 镰刀需(Fusarium) 黄孢原毛平革黄 (Phanerochaete) 毛霜菌(Mucor) 曲雷菌(Aspergillus) 图1.9真菌 的酵母还能用于微生物排泄物的 类却更为丰富,而且有些种类甚至 真菌(fungi)在自然循环,尤 探别(见第六章和第十章)。 能靠纤维素(里氏木客,Tricho- 其在分解过程中,具有极其重要 客萄(mold fungi)是子食菌 derma reesei)和木质素(黄袍原毛 的作用。运今为止,大约有7万种 口(Ascomycota)中的一贡,子袁 平羊菌,Phanerochaete chrysospo 真菌已被分门别是。像所有真菌 菌是真菌大家族中种美最多的一 rim)生活(见第六章)。 一样,酵垂(yeast)属于真核生 类,包括两万种真菌。与球状酵母 曲车属菌(Aspergill)与青 物,具有贮存全部遣传信息的细 不同,子数菌又细又长,而且地大 客属菌(Penicillium)都常被用于 胞核。 多数都是绝对需乳的。雾菌艳子 发酵,尤其是酶解过程(酶解大 酵母的种类有很多,我们要 是通过无性繁殖的方式,由菌丝 麦、小麦、黄豆等),黑曲客首 把野生酵母与大规模人工培养的 体如胞核一分为二形成的。葡生 (Aspergillus niger)还会产生柠檬 琼脂培养基上的黑曲客(Aper 酵母区分开来,比如啤酒酵#、面 体逐渐延仲,就形成了孢子囊。这 酸(见第四章)。其他种夹的青年 gillus niger)。 包酵量和饲科酵母。产阮组丝砖 样,成熟的弛子就会随空气的流 菌则用于生产特殊种类的奶酪, 母(Candida utilis)通常生长在造 动而四处漂走,当找到追于生长 p软质白霉乳酪(Camembert)浅 纸厂排出的富舍亚硫酸盐的木菜 的环境时,就会形成新的菌丝体, 者洛克福羊乳千酪(Roquefort), 废物中。麦芽糖复丝酵母 除此之外,真菌还可以根据形态 (Candida maltosa)是一种生长在 和柄色进行更细的分类。 真菌生产的这粉酶类和爱白 酶类还可以用作酶制剂的商业化 原油中的真菌,利用原油中的烷 这些菌拉体的外形美不多,都 生产(见第二幸)。 烃产生饲料蛋白,皮状址弛酵母 是一团线状的菌丝。在大规模培养 (Trichosporon cutaneum)能够分 的培养罐中,很多人工酵章部只能 拟内孢客(Endomycopsis)与 解排放物中对其他真菌有毒害作 产生团被的菌丝,却不能生成孢 毛霉(Mcor)也可用于大规模生 用的酚,丝袍酵母(Trichosporon) 子.与一校的野生酵母一样,人工 产酶制剂,而缘刀首(Fusarium)则 和一-种叫微Arxula adeninivorans 酵母也需要营养物质,但雪要的种 可为人类提供蛋白质(见第六章)。 产生青雾素的零菌一点青客 (Penicillium notatum)
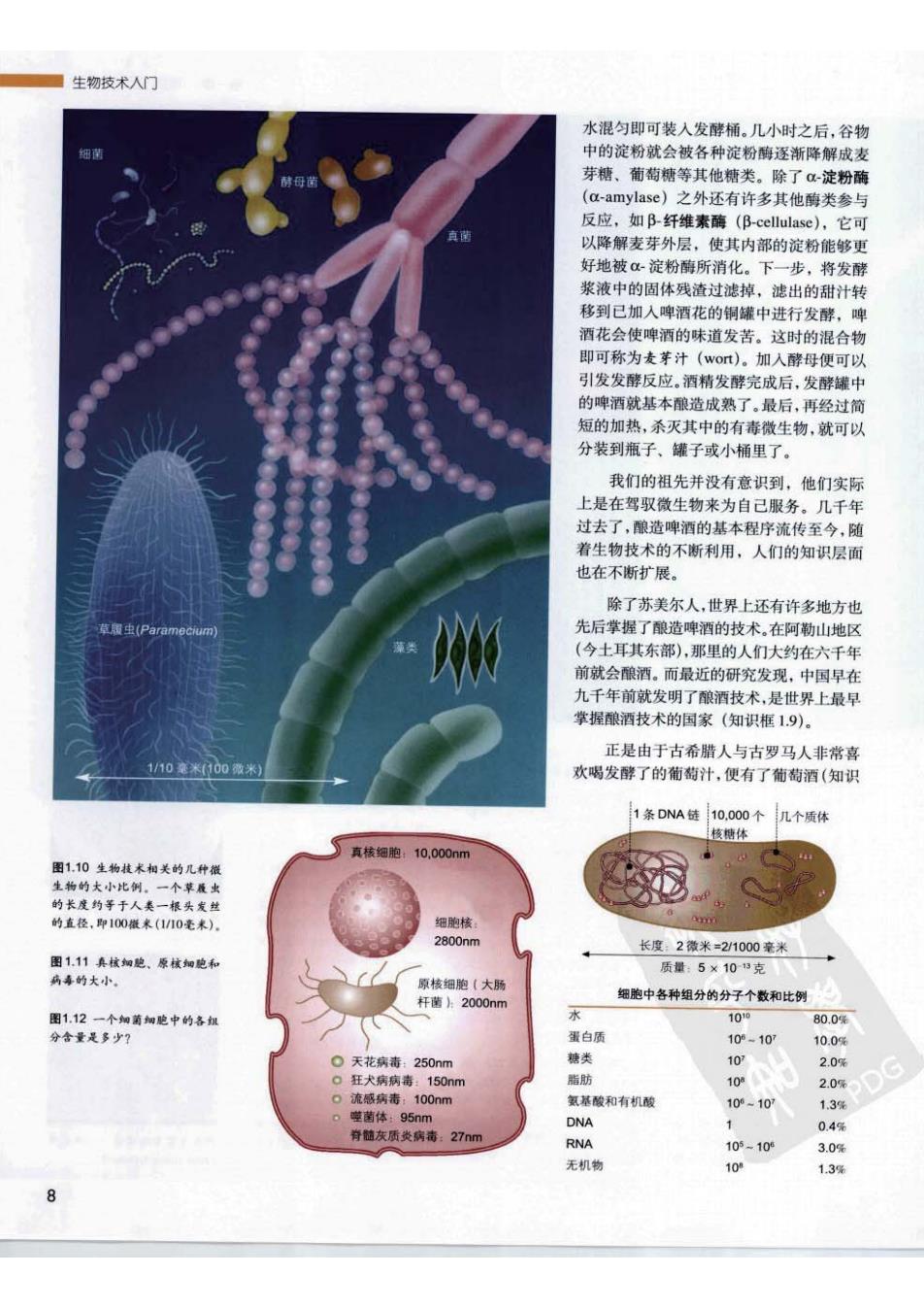
生物技术人门 水混匀即可装入发酵桶。几小时之后,谷物 细带 中的淀粉就会被各种淀粉酶逐渐降解成麦 芽糖、葡萄糖等其他糖类。除了-淀粉酶 (a-amylase)之外还有许多其他酶类参与 反应,如B纤维素酶(B-cellulase),它可 真 以降解麦芽外层,使其内部的淀粉能够更 好地被淀粉酶所消化。下一步,将发酵 浆液中的固体残渣过滤掉,滤出的甜汁转 移到已加入啤酒花的铜罐中进行发酵,啤 酒花会使啤酒的味道发苦。这时的混合物 即可称为麦芽汁(wot)。加入酵母便可以 引发发酵反应。酒精发酵完成后,发酵罐中 的啤酒就基本酿造成熟了,最后,再经过简 短的加热,杀灭其中的有毒微生物,就可以 分装到瓶子、罐子或小桶里了。 我们的祖先并没有意识到,他们实际 上是在驾驭微生物来为自己服务。几千年 过去了,酿造啤酒的基本程序流传至今,随 着生物技术的不断利用,人们的知识层面 也在不断扩展。 除了苏美尔人,世界上还有许多地方也 草银(Paramcum 先后掌握了酿造啤酒的技术。在阿勒山地区 (今土耳其东部),那里的人们大约在六千年 前就会酿酒。而最近的研究发现,中国早在 九千年前就发明了酿酒技术,是世界上最早 掌握酿酒技术的国家(知识框1.9)。 正是由于古希腊人与古罗马人非常喜 1/10毫米(300微米 欢喝发酵了的葡萄汁,便有了葡萄酒(知识 1条DNA链 10.000个 几个质体 核糖体 真核细胞:10.000nm 图110生物枝术相关的几种橙 生物的大小比例。一个草履出 的长度约等于人类一根头发丝 的直径,即100橙米(1/10毫米)。 细胞核 2800nm 长度2微米=2/1000毫米 图1.11真核如胞、原核细胞和 质量:5×103克 病喜的大小。 原核细胞(大肠 杆菌片2000nm 细胞中各种组分的分子个数和比例 图1.12一个细菌细胞中的各级 10° 80.0% 分含量是多少? 蛋白质 109-107 10.0% 。天花病毒:250nm 糖类 102 2.0% 0狂犬病病毒:150nm 脂肪 10 4 2.0% 0流感病毒:100nm 氢基酸和有机酸 100-107 1.3% 噬菌体:95nm DNA 肯随灰质炎病毒:27nm 0.4% RNA 105-106 3.0% 无机物 10 1.3% 8

第一章啤酒,面包,奶酪 知识框1.3葡萄酒与烈性酒 萄酒,酒精浓度大多在10%-15%。 为32%,在发酵加工过程中还常常会添 葡萄酒可以通过颜色(主要是红色 加少量的芳香物质,如洋茴香、茴香、香 和白色)、产地及葡萄的品种进行分类。 菜或者杜松。谷物烈酒的原材料通常选 为了使葡萄酒残留一丝甜味,可以中断 用黑麦、小麦、养麦、燕麦或大麦。制造 发酵过程,加入些许辅助材料,如新榨 威士忌(酒精含量至少为40%)时,为 的葡萄汁。而干葡萄酒(无甜味的葡萄 了使其产生薰香味,会在烘麦芽时往烘 酒)含糖量最多为9克/升,半干葡萄酒 炉中加入一些泥炭。伏特加(酒精含量为 最多为18克/升,超过18克/升的就是 40%-60%)是用黑麦、马铃薯及其他淀 甜葡萄酒了。还有一种加强型葡萄酒, 粉类植物制成的,发酵过程中,麦芽浆需 其中加人了糖和酒精,有时甚至是香 要反复不断地蒸馏。在加工金酒(即杜松 草,但加入这些材料后不再需要后续的 子酒)的过程中,要在麦芽浆中加入杜松 发酵过程。 子或醇制杜松子的提取物。 果汁白兰地(酒精含量至少为38%) 般不需要添加任何糖、乙醇或色素,直 接由水果发酵制成,品种包括:苹果白兰 地、果渣葡萄酒,而像樱桃酒、梅子酒、 杏酒、桃子酒这类酒则是用新鲜水果添 在葡萄酒的制备过程中,第一步就 加酒精制成。白兰地必须由酒或者发酵 是将葡萄放入碾磨机中榨出葡萄汁。制 的水果汁蒸馏提取制成。 作白葡萄酒时,必须尽快捣磨葡萄,将 葡萄汁与果梗、果皮、果籽等榨汁剩下 南非Cape镇附近Stellenbosch的葡首国。 法国干邑葡萄酒是一种蒸馏酒,得 的果渣分离。如果在榨取过程中加入果 香槟和其他汽酒 名于它的发源地—法国的干邑镇 胶酶(pectinase,见第二章),则有利于 (Cognac)。除此之外,法国西南部的热 得到更多更澄清的葡萄汁。制作红葡萄 香槟及其他汽酒需要进行两次发 尔省境内、干邑南部的雅邑镇,以及其他 酒时,捣磨时间比较长(45天),但不 酵,二氧化碳是最后被封入瓶中的。第 国家如德国、希腊、意大利、西班牙等地 需要将葡萄汁与果渣分离,直接将捣磨 二次发酵开始时,葡萄酒除了与一些特 也生产类似的葡萄酒白兰地。朗姆酒是 得到的葡萄浆发酵即可,因为葡萄皮中 殊的香槟酵母混合加入瓶中外,还需添 用甘蔗榨汁发酵蒸馏得到的,其酒精含 的花色甙会在发酵过程中被慢慢溶解, 加一些糖浆。在之后的几个月里,将瓶 量为38%。亚力酒则是用稻子或椰子花 使葡萄浆的颜色变红。 子逐渐倾斜,使酒液聚集到瓶子的上 制成。 部,而酵母及其他沉淀就会聚集在瓶颈 发酵可以直接利用依附在葡萄外皮 部位。这时,将瓶塞拔去便可除去沉淀 利口酒多采用芳香植物及药用植物 上的酵母菌,也可以接入纯的酿酒酵母 物,随后加入一些糖浆和白兰地,然后 的根.茎、叶、果和果浆作为添加料。宾 培养物(yeast cultures)。发酵过程中会 换用缠线的软木塞塞住瓶口。这样产生 治酒(punch,源自北印度语punsha,意 产生许多泡沫。这种带泡沫的混浊液在 的稳定且优质的香槟会在启塞之后自行 思是“五个“)是一种热饮酒,由五种原 德国南部许多地区都十分受欢迎,在秋 产生适量的气泡,而不像其他的汽酒是 料制成:酒精、香料、柠檬汁、糖、水或 季配上洋葱蛋挞,简直就是一道美味。 将二氧化碳压人瓶中。 茶水,鸡尾酒是一种开胃酒,起源于美国 独立战争时期,它将各种不同的酒倒在 约4~8天后,发酵过程已基本完成, 烈性酒 起而不混合,各种酒的颜色层层堆叠, 其中的糖分也基本消耗殆尽,但蛋白质 烈性酒包括白兰地(brandy)、利口 宛如美丽多彩的羽毛。 与果胶却没有被溶解,因此需要将两者 沉淀、与葡萄酒分离。之后的十二个月 酒((liqueur)、宾治酒(punch)以及鸡尾 里,将葡萄酒贮藏在阴冷的地窖中,残 酒(cocktails)。法国干邑葡萄酒(Cog 余的糖分会继续进行缓慢发酵,并产生 nac)、阿马涅克白兰地(Armagnac)、朗 沉淀。这时的葡萄酒就能散发出浓郁的 姆酒(rum)、亚力酒(arrack)、威士忌 酒香和独特的风味。发酵结束后,将葡 (whisky)以及果汁酒(fruit schnapps)都 萄酒装人预先用硫磺处理过的木桶中, 是众所周知的烈性酒。发酵过程中的副 塞紧桶塞,待葡萄酒继续酿熟。在最后 产物(酯类、高级醇、醛类、酸等等)会 的酿熟阶段,可以定时地打开塞子透透 在蒸馏时部分地保存下来,这样可以优 在南非Stellen 空气,硫磺的氧化作用可以产生二氧化 化酒的口感。一般来说,淀粉发酵则不 bosch的一个酿 硫,起到抑制细菌的作用,以提高葡萄 会产生大量的高级醇。 酒厂中,作者想 要钻进空的发酵 酒的贮存稳定性。这种方法酿造出的葡 通常情况下,烈酒的乙醇含量至少 罐一探究竞
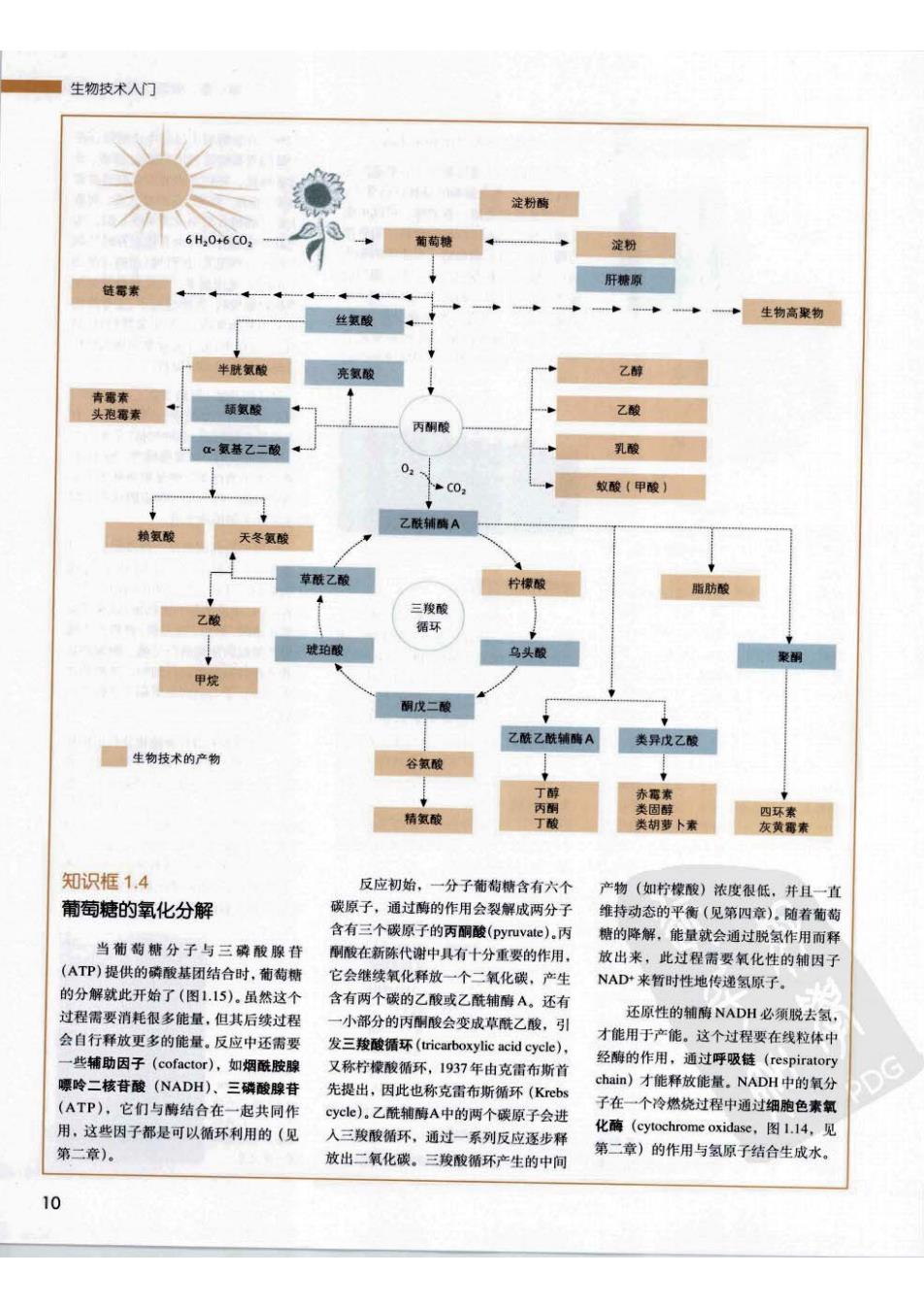
生物技术人门 淀粉酶 6H20+6C02 葡萄糖 淀粉 肝糖原 链霉素 丝氨酸 生物高聚物 半胱氨酸 亮氢酸 乙醇 青需素 头泡霉素 颉氨酸 乙酸 丙酮酸 c-氨基乙二酸 乳酸 02 C02 蚁酸(甲酸) 乙酰辅酶A 赖氨酸 天冬氨酸 草酰乙酸 柠檬酸 脂肪酸 三羧酸 乙酸 循环 琥珀酸 乌头酸 聚钢 甲烷 酮戊二酸 乙酰乙酰辅酶A 类异戊乙酸 生物技术的产物 谷氢酸 丁醇 赤霭素 丙酮 类固醇 精氨酸 四环素 丁酸 类胡萝卜素 灰黄霉素 知识框1.4 反应初始,一分子葡糊糖含有六个 产物(如柠檬酸)浓度很低,并且一直 葡萄糖的氧化分解 碳原子,通过酶的作用会裂解成两分子 维持动态的平衡(见第四章)。随着葡萄 含有三个碳原子的丙酮酸(pyruvate).丙 糖的降解,能量就会通过脱氢作用而释 当葡萄糖分子与三磷酸腺苷 酮酸在新陈代谢中具有十分重要的作用, 放出来,此过程需要氧化性的辅因子 (ATP)提供的磷酸基团结合时,葡萄糖 它会继续氧化释放一个二氧化碳,产生 NAD来暂时性地传递氢原子。 的分解就此开始了(图1.15)。虽然这个 含有两个碳的乙酸或乙酰辅醇A。还有 过程需要消耗很多能量,但其后续过程 一小部分的丙酮酸会变成草酰乙酸,引 还原性的辅酶NADH必须脱去氢, 会自行释放更多的能量。反应中还需要 发三羧酸循环((tricarboxylic acid cycle), 才能用于产能。这个过程要在线粒体中 一些辅助因子(cofactor),如烟酰胺腺 又称柠檬酸循环,1937年由克雷布斯首 经酶的作用,通过呼吸链(respiratory 嘌呤二核苷酸(NADH)、三磷酸腺苷 DG 先提出,因此也称克雷布斯循环(Krebs chain)才能释放能量。NADH中的氧分 (ATP),它们与酶结合在一起共同作 cycle)。乙酰辅酶A中的两个碳原子会进 子在一个冷燃烧过程中通过细胞色素氧 用,这些因子都是可以循环利用的(见 入三羧酸循环,通过一系列反应逐步释 化酶(cytochrome oxidase,图l.14,见 第二章)。 放出二氧化碳。三羧酸循环产生的中间 第二章)的作用与氢原子结合生成水。 10

第一章啤酒,面包,奶酪 框13),随后,也是古罗马人将这项酿酒技 料,另一方面,还要为生物合成及其他耗能 被还原的辅酶因子 术传到了现在的德国和英国。 反应提供能量。所需的这些能量都是通过 在非洲,当地人用裂殖酵母(Schi 摄取营养物质而获得的一营养物质首先 NAD zo.saccharomyces pombe)酿造出非洲酒 分解成较小的颗粒,再降解成小分子物质, (pombe),中亚的人们则用马奶为原料 这些小分子物质便组成了细胞构建所需的 制成奶酒(kumys),当地人还将奶酒装 原材料,如葡萄糖、氨基酸、核苷酸(嘌呤1 入皮质的酒囊中随身携带,日本人用稻 嘧啶碱基及磷酸核糖)、有机酸、脂类等等。 米酿制出米酒(sake):俄国人用乳酸杆菌 它们都是大分子物质如蛋白质、核酸、多糖 (Lactobacillus)和米曲霉(Aspergillus 及细胞壁的组成物。 乙醇 oryzae)的糖化作用酿造出了淡啤酒格瓦斯 碳水化合物是光合作用的主要产物, 脱氢酶 (kvass)。 这为大多数有机体提供了营养基础,因此, 关于葡萄酒的酿造技术(知识框13), 葡萄糖(glucose)就成为许多代谢过程的 几千年来并没有什么变化:首先将新鲜的 起始物质。正是这些降解葡萄糖的代谢过 红葡萄或白葡萄采摘下来,搅碎并压榨出 程,引发了生物技术生产中许多重要的途 汁液来,将榨出的葡萄汁放入密封的罐子 径。 被氧化的 中发酵即可。在古代,人们用木桶发酵,现 ■1.5酒精发酵:酵母生存的应急 辅酶因子 如今则使用容量高达25万升的大金属罐, 措施 NAD+ 全世界葡萄酒年产量更高达500亿升。 通过生物化学的基础知识可以知道, ■1.4万物生长靠太阳 酵母是一类兼性好氧的微生物—它们既 乙醇 太阳以电磁波辐射的形式向四周发出 可以进行有氧呼吸,也可以进行无氧发酵。 图1.13酵母中的乙醇脱氢酶将 巨大能量。太阳每天向地球释放出高达 处于有氧环境下,酵母可以很好地生 乙醛转化为C醇。在生物分析法 中(见第十章),这个可逆反应常 4000万亿千瓦小时(度)的能量,仅30分 长,它们通过呼吸将糖类转化成二氧化碳 用来测定血液中的酒精含量水平, 钟向地球投射的太阳能总量就足够全球生 和水,并释放出能量,用于各种生命活动, 物一年能量的需求。然而在这些能量中, 如细胞生长和分裂。 只有三千分之一的光能被绿色植物通过光 当空气供应不足时,酵母就会将新陈 氧分子 合作用(photosynthesis)转化成化学能。 代谢转换成为无氧应急模式(anaerobic 绿色植物叶绿体中的能荷光子将水分 emergency mode),尽管无氧发酵的产能效 还原当量 解成氢原子和氧原子(每年1千亿吨)。只 率较低,但却能使酵母在缺氧的逆境中得 有释放到大气层中的氧分子才能被那些有 以存活。1861年,路易斯·巴斯德(Louis 氧呼吸的有机体所利用,而光合作用产生 Pasteur,1822-1895)发现:在缺氧条件下 的氢可以暂时与碳结合成为碳水化合物 酵母消耗的葡萄糖分子数远远大于有氧条 (carbohydrate)。碳水化合物是光合作用的 件下对葡萄糖的使用,这就是众所周知的 主要产物,也是大多数生物所能利用的主 “巴斯德效应”(Pasteur effect),即由于无 要能源。生物可以使碳水化合物中的氢与 氧发酵的产能率低,缺氧条件下酵母会消 碳分离,通过生化爆鸣气反应(biochemical 耗比有氧条件更多的葡萄糖来补偿能量的 detonating gas reaction)释放氢,同时通过 损耗。 呼吸链产生能量。而这一切都不得不归功 缺氧时,由于呼吸链不能彻底进行, 于这个巨大的天然能源一太阳! 产生的NADH+H不能进入三羧酸循环而 不论处于生长期还是休眠期,细胞都需 只能逐渐停止在丙酮酸环节,细胞则利用 图1.14细胞色素乘化酶是一种系 要源源不断的能量供应。从营养物质吸收的 NADH+H+、经乙醛脱氢酶的作用将丙酮 与细胞呼吸的醉,最早由奥托·沃 能量又通过细胞新陈代谢(metabolism)中 伯格(Otto Warburg)发现.在o 酸转化成乙醛并释放出二氧化碳, 胞畔吸这种“冷然烧”过程中,此 的各个途径而逐级消耗殆尽。这些代谢途 NADH+H+也随之脱氢转化成NAD+(图 酶能将夏转化为水并同时释放能 径一方面能够为细胞生长提供所需的原材 1.13)。换句话说就是,葡萄糖通过不完全 11