
蛋白质, 直径为20-25nm 。真核细胞的核蛋白体比原核细胞的大,其沉降系数一般为80S,它由60S和40S的 组成。细胞质核蛋白体有的呈游离状态,有的和内质网及核膜结合。线粒体核蛋白体存在于线粒休内膜的 间,但沉降网 是存在于细胞质中折叠的膜系统。 典型的内质网是成对的平 的腔分隔形成封闭的 由狭窄 由质上 于常被核蛋白体附若形成粗糙内 在丝的顶端细胞中常可见到 面的内质网称为光滑型内质网。内质网沟通若细胞的各个部分, 与细而细地 线休 都右群系内历 是细胞中各种物质运转的一种循环系统,同时内质网还供给细胞质中所有细胞器的膜。 ③线粒体(@itochondria)线粒体是含有DNA的细胞器。它具有双层膜,内层较厚,常向内延伸形成不同数量 和形状的脊。线粒体的形态、数量和分布常因真菌的种类和发育阶段而异。线粒体是氧化磷酸化作用和ATP形成的场 所。其内膜上有细胞色素、NDH脱氢酶、琥珀酸脱氢酶和ATP磷酸化酶,此外三羧酸循环的酶类、核糖核蛋白体, 蛋白质合成酶和DNA,以及脂肪酸氧化作用的酶也都在内膜上。外膜上也有多种酶,如脂类代谢的酶类等。总之,线 体是 细跑的“动力房 均 布 的 往往上内 少数几种真菌中发 是某些真菌菌丝细胞中由单层膜包裹的细胞器 膜之间。形态呈管状、囊 状、球状、卵圆形或作多层折叠的膜,内含泡状物或颗粒状物。有些边体与细胞膜连接在一起。边体分泌水解酶或与 细胞壁的形成有关。 ⑥溶酶体 1s。s0m)是一种由单层膜句鬼、内含多种酸性水解酶的小成形、蜜随状细胞器。含有多种酸性 水解酶,主要功能是细胞内的消化、维持细胞营养及防止外米微生物或异体物质侵袭的作用。 ⑦微 其功能 (microbody) 是一种由单层膜包裹、与溶酶体相似的小球形细胞器,主要含氧化酶和过氧化氢酶, 胞免受H 202每 并能氧化分角 vacuole 由单位膜分隔 形色 天小受细胞年龄 和生到 状态而变化 ·般在老龄细胞中液泡 以 还有溶酶体的助 可以把白水解细离 4、细胞核 真菌细胞核通常为椭圆形,直径一般为23μⅢ 能通过菌丝隔膜上的小孔在菌丝中很快地移动。用相差显微 镜观察真菌活细胞,可观察到被一层均匀的核质包围的中心稠密区,即核仁。核仁除DNM外还含有RNA,但RNA在 细胞核分裂时消失。核膜一般为两层,厚820m。膜上有小孔,以利核内外物质交流。核膜孔径大小差异很大,孔 的数量随菌龄而增大。核膜的外膜常有核蛋白附着。真菌核膜在核的分裂过程中一直存在,这与其他高等生物不同。 5、鞭毛图2.2真菌鞭毛横切面示意图 白毛处 长度为 中央微苦 毛与细菌的鞭毛 动能上虽相同,但 在构造、运动机理和所耗能源形式等 方面金 微管连批置白 徽管二联体 有显著差别」 动力蛋白时 真菌鞭毛由鞭杆(shaft)和基体 一硬毛外膜 (basel body)组成。鞭杆的横切面呈 央微管桥 “9+2”型,即中心有一对包在中央鞘内 的相互平行的中央微管,其外围绕一圈(9个)微管 成 话性 联体,整个鞭杆由细胞质膜包裹。 联体是由亚 鞭毛轴丝中: 丝。和B组成的, 根原纤维形成。 B仅体的 TP醇活性 臂长约 1管伸出 亚 成相邻 联体微管之间连接 在二联体之间有连丝连接。此外 还从亚丝A向中央 对微管发出辐射状的连丝,称为放射辐。放射辐的辐头 周控微管的起始和延伸 是自由的, 不与中中髓相连 基体又称生毛体或动体,呈短杆状,直径约 促讲环中和微管中微管蛋自的聚 蛋白激南 尚管蛋白亚单位的威融化 120^170nm,长为200^500nm,横切面观察,外围有9 个 联体,中央没有微管和鞘,为9+0型(图2.2) 表22某些已知与微管或微管蛋白相互作用的蛋白质 ssociated protein)共同组成,它们与运动的 微管聚合和解聚的调节或微管和其它细胞组成 一在三联体的侧上存在的P解,称为动力蛋白同的等相天水解AP,放 放出能量,供鞭毛运动所需
蛋白质,直径为 20-25nm 。真核细胞的核蛋白体比原核细胞的大,其沉降系数一般为 80S ,它由 60 S 和 40 S 的 2 个小亚基组成。细胞质核蛋白体有的呈游离状态,有的和内质网及核膜结合。线粒体核蛋白体存在于线粒体内膜的嵴 间,但沉降系数为 70 S 。 ② 内质网 (endoplasmic reticulum) 是存在于细胞质中折叠的膜系统。典型的内质网是成对的平行膜,由狭窄 的腔分隔形成封闭的管道系统,有时是分枝的管道。内质网的主要成分是脂蛋白,但游离蛋白和其他物质有时也合并 到内质网上,且时常被核蛋白体附着形成粗糙型内质网,这在菌丝的顶端细胞中常可见到。没有被核蛋白体附着在上 面的内质网称为光滑型内质网。内质网沟通着细胞的各个部分,它与细胞质膜、细胞核、线粒体等都有联系。内质网 是细胞中各种物质运转的一种循环系统,同时内质网还供给细胞质中所有细胞器的膜。 ③ 线粒体 (mitochondria) 线粒体是含有 DNA 的细胞器。它具有双层膜,内层较厚,常向内延伸形成不同数量 和形状的脊。线粒体的形态、数量和分布常因真菌的种类和发育阶段而异。线粒体是氧化磷酸化作用和 ATP 形成的场 所。其内膜上有细胞色素、 NADH 脱氢酶、琥珀酸脱氢酶和 ATP 磷酸化酶,此外三羧酸循环的酶类、核糖核蛋白体、 蛋白质合成酶和 DNA ,以及脂肪酸氧化作用的酶也都在内膜上。外膜上也有多种酶,如脂类代谢的酶类等。总之,线 粒体是酶的载体,细胞的“动力房”。 ④ 高尔基体 (dictyosome ,或 Golgi body) 真菌的高尔基体在细胞中大多呈网状,少数为鳞片状、颗粒状或杆 状,均匀分布于核的周围,往往与内质网相连。高尔基体与细胞的分泌机能有关,是凝集某些酶原颗粒 ( 如消化酶原 ) 的场所,且与细胞膜的形成以及碳水化合物的合成有关。目前高尔基体仅在少数几种真菌中发现。 ⑤ 边体 (lomasome) 是某些真菌菌丝细胞中由单层膜包裹的细胞器,位于细胞壁和细胞膜之间。形态呈管状、囊 状、球状、卵圆形或作多层折叠的膜,内含泡状物或颗粒状物。有些边体与细胞膜连接在一起。边体分泌水解酶或与 细胞壁的形成有关。 ⑥ 溶酶体 ( lysosome ) 是一种由单层膜包裹、内含多种酸性水解酶的小球形、囊膜状细胞器,含有多种酸性 水解酶,主要功能是细胞内的消化、维持细胞营养及防止外来微生物或异体物质侵袭的作用。 ⑦ 微体 ( microbody ) 是一种由单层膜包裹、与溶酶体相似的小球形细胞器,主要含氧化酶和过氧化氢酶, 其功能可使细胞免受 H 2 O 2 毒害 ,并能氧化分解脂肪酸等。 ⑧ 液泡 ( vacuole ) 由单位膜分隔,其形态、大小受细胞年龄和生理状态而变化,一般在老龄细胞中液泡大 而明显。真菌的液泡中主要含有糖原、脂肪和多磷酸盐等贮藏物,精氨酸、鸟氨酸和谷氨酰胺等碱性氨基酸,以及蛋 白酶、酸性和碱性磷酸酯酶、纤维素酶和核酸酶等各种酶类。液泡不仅有维持细胞渗透压、贮存营养物等功能,而且 还有溶酶体的功能,因为它可以把蛋白酶等水解酶与细胞隔离,防止细胞损伤。 4 、细胞核 真菌细胞核通常为椭圆形,直径一般为 2~3 μ m ,能通过菌丝隔膜上的小孔在菌丝中很快地移动。用相差显微 镜观察真菌活细胞,可观察到被一层均匀的核质包围的中心稠密区,即核仁。核仁除 DNA 外还含有 RNA ,但 RNA 在 细胞核分裂时消失。核膜一般为两层,厚 8~20nm 。膜上有小孔,以利核内外物质交流。核膜孔径大小差异很大,孔 的数量随菌龄而增大。核膜的外膜常有核蛋白附着。真菌核膜在核的分裂过程中一直存在,这与其他高等生物不同。 5 、鞭毛 图 2.2 真菌鞭毛横切面示意图 在低等水生真菌游动孢子和配子表面 有能产生运动的细胞附属器即鞭毛,单极生 或双极生,长度为 20~2000 μ m 。真菌的 鞭毛与细菌的鞭毛在运动功能上虽相同,但 在构造、运动机理和所耗能源形式等方面都 有显著差别。 真菌鞭毛由鞭杆( shaft )和基体 ( basel body )组成。鞭杆的横切面呈 “ 9+2 ”型,即中心有一对包在中央鞘内 的相互平行的中央微管,其外围绕一圈( 9 个)微管二 联体,整个鞭杆由细胞质膜包裹。每一个二联体是由亚 丝 A 和 B 组成的,亚丝 A 由 13 根原纤维形成。亚丝 B 仅 10 根原纤维,其中有 3 根是与亚丝 A 合用的。 从二联体的 A 管伸出二条侧臂,对着相邻二联体的亚丝 B,臂长约 30nm,宽 9nm,两臂之间的间隔为 16-22nm 。 在二联体之间有连丝连接。此外,还从亚丝 A 向中央一 对微管发出辐射状的连丝,称为放射辐。放射辐的辐头 是自由的,不与中央鞘相连。 基体又称生毛体或动体,呈短杆状,直径约 120~170nm ,长为 200~500nm ,横切面观察,外围有 9 个三联体,中央没有微管和鞘,为 9+0 型 ( 图 2.2) 。 表 2 -2 某些已知与微管或微管蛋白相互作用的蛋白质 真核微生物鞭毛微管主要由微管蛋白和微管相关蛋白( Microtuble-associated protein )共同组成,它们与运动的 产生、微管聚合和解聚的调节或微管和其它细胞组成之间的连接等相关(表 2-2 )。 在二联体的侧臂上存在的 ATP 酶,称为动力蛋白( dynein ),它可以水解 ATP ,放出能量,供鞭毛运动所需。 成分 活性 在鞭毛轴丝中: 动力蛋白 1 动力蛋白 2 连接蛋白 ATP 酶活性 产生运动 形成相邻二联体微管之间连接 在细胞质微管中: 高分子量蛋白质 tau 蛋白激酶 MAP1 MAP2 调控微管的起始和延伸 促进环中和微管中微管蛋白的聚合 微管蛋白亚单位的磷酸化
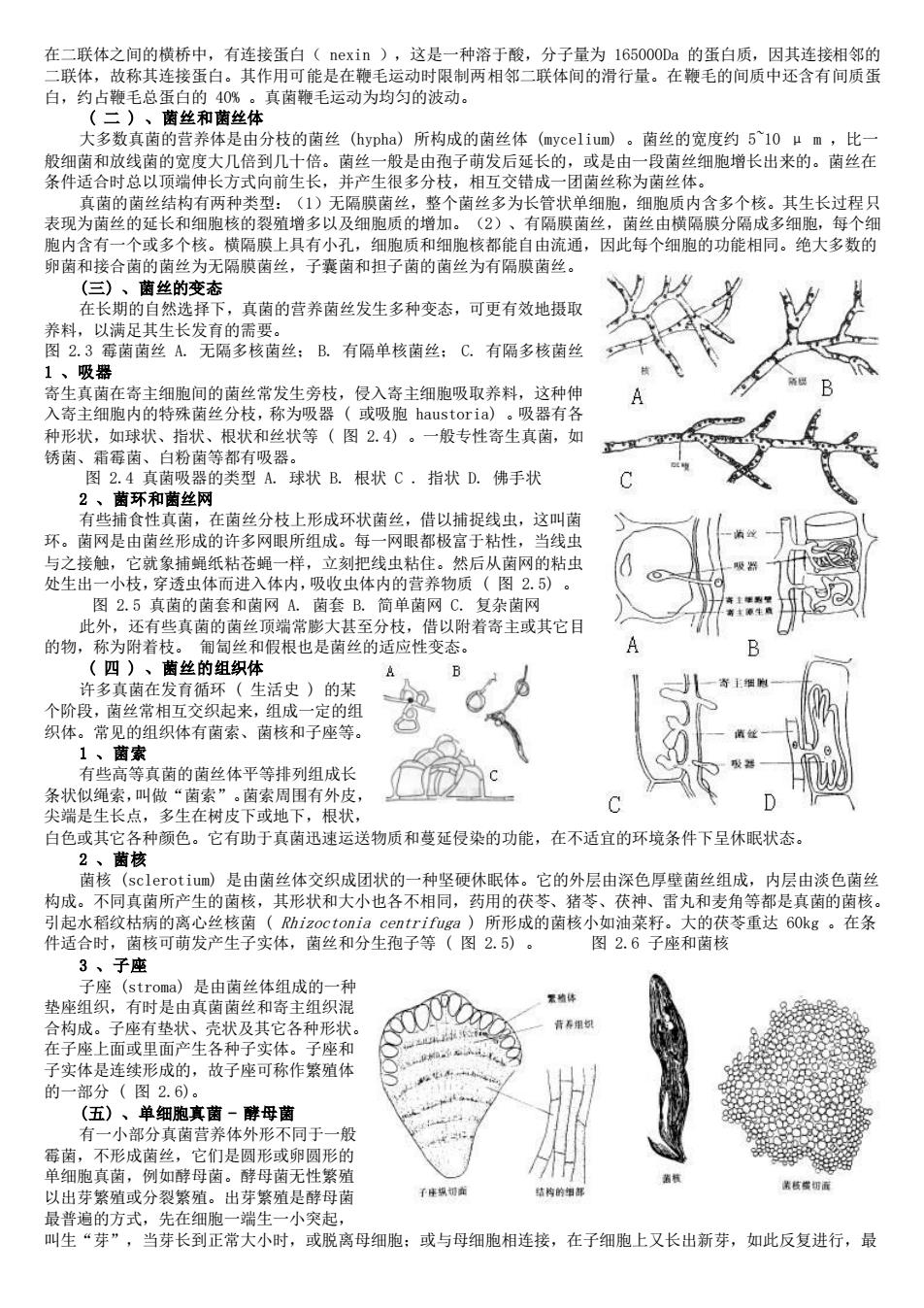
在二联体之间的横桥中 有连接蛋白(nexin),这是一种溶于酸,分子量为165000Da的蛋白质,因其连接相邻的 二联体,故称其连接蛋白。其作用可能 是在鞭 毛运动时限制两相邻二联体间的滑行量。在鞭毛的间质中还含有间质蛋 白,约占鞭毛总蛋 0 真菌鞭毛运动为均匀的波动。 分枝的菌丝(ypha)所构成的菌丝 菌丝的宽 510 细胞增长出来的。 向 面产生很多分枝 错成 菌丝布 (1)无隔膜菌丝,整个菌丝多为长管状单细胞,细胞质内含多个核。其生长过程只 表现为茵丝的延长和细胞核的裂殖增多以及细胞质的增加。 (2)、右隔隙菌丝,茵丝由惜隔膜分隔成多细胞。每个细 胞内含有 个或多个核。横隔膜上具有小孔,细胞质和细胞核都能自由流通,因此每个细胞的功能相同。绝大多数的 卵菌和接合菌的菌丝为无隔膜菌丝,子囊菌和担子菌的菌丝为有隔膜南丝。 (三)、菌丝的变态 在长期的目然选下 ,真菌的营养菌丝发生多种变态,可更有效地摄取 C.有隔多核菌丝 寄生真茵在寄主细胞间的菌丝常发生旁枝,侵入寄主细胞吸取养料, 这种伸 入寄主细胞内的特殊南丝分枝,称为吸器(或吸胞hau 种形状,如球状、指状、根状和丝状等(图24)。 一般专性寄生真菌,如 锈菌、霜莓菌、白粉菌等都有吸器。 图2.4真茵吸器的类型A.球状B.根状C,指状D.佛手状 2、菌环和菌丝网 捕食性真菌,在菌丝分枝上形成环状菌丝,借以捕捉线虫,这叫菌 环。菌网是由菌丝形成的许多网眼所组成。每一网眼都极富于粘性, 白线里 样 虫 然后从 网的粘虫 复杂 还有些真茵的菌 昔以附者寄主或其它目 的物, 敌为附若 匍匐丝和假根也是菌丝的适应性变态。 (四)、 菌丝的组织体 许多直菌在发有循环(生活中)的其 个阶段,菌丝常相互交织起来,组成一定的组 织体。常见的组织体有菌索、菌核和子座等。 1、菌索 高等真菌的菌丝体平等排列组成长 条状绳素叫做生 白色或其各种颜色。它有助子其寄迅速运送物质和蔓延侵染的功能,在不适宜的环境条件下呈休状态 萌核 )是由茵丝体交织成团状的一种坚便休眼体。它的外层由深色厚壁菌丝组成,内层由淡色菌丝 构成。不同真菌所产生的菌核,其形状和大小也各不相同,药用的茯苓、猪苓、茯神、雷丸和麦角等都是真菌的南核, 引起水稻纹枯病的离心丝核菌(Rhizoctonia centrifuga)所形成的菌核小如油莱籽。大的茯苓重达6Okg。在条 件适合时,南核可萌发产生子实体,南丝和分生孢子等(图2.5)。 图2.6子座和南核 3、子座 子座(stroma)是由菌丝体组成的一种 垫座组织,有时是由真菌菌丝和寄主组织 植体 面产生各种之 各种形获 本是连续形 的 实体分(国26 故 可称作繁殖 (五)、单细跑直黄一母菌 有一小部分真菌营养体外形不同于一般 莓菌 不形成菌丝 它们是圆形或卵圆形的 单细胞真菌,例如酵母菌。酵母菌无性繁殖 以出芽紧殖或分裂紧咀。出芽紧殖是酵母陶 最普遍的方式,先在细胞一端生一小突起, 叫生“芽”,当芽长到正常大小时,或脱离母细胞:或与母细胞相连接,在子细胞上又长出新芽,如此反复进行,最
在二联体之间的横桥中,有连接蛋白( nexin ),这是一种溶于酸,分子量为 165000Da 的蛋白质,因其连接相邻的 二联体,故称其连接蛋白。其作用可能是在鞭毛运动时限制两相邻二联体间的滑行量。在鞭毛的间质中还含有间质蛋 白,约占鞭毛总蛋白的 40% 。真菌鞭毛运动为均匀的波动。 ( 二 ) 、菌丝和菌丝体 大多数真菌的营养体是由分枝的菌丝 (hypha) 所构成的菌丝体 (mycelium) 。菌丝的宽度约 5~10 μ m ,比一 般细菌和放线菌的宽度大几倍到几十倍。菌丝一般是由孢子萌发后延长的,或是由一段菌丝细胞增长出来的。菌丝在 条件适合时总以顶端伸长方式向前生长,并产生很多分枝,相互交错成一团菌丝称为菌丝体。 真菌的菌丝结构有两种类型:(1)无隔膜菌丝,整个菌丝多为长管状单细胞,细胞质内含多个核。其生长过程只 表现为菌丝的延长和细胞核的裂殖增多以及细胞质的增加。(2)、有隔膜菌丝,菌丝由横隔膜分隔成多细胞,每个细 胞内含有一个或多个核。横隔膜上具有小孔,细胞质和细胞核都能自由流通,因此每个细胞的功能相同。绝大多数的 卵菌和接合菌的菌丝为无隔膜菌丝,子囊菌和担子菌的菌丝为有隔膜菌丝。 (三) 、菌丝的变态 在长期的自然选择下,真菌的营养菌丝发生多种变态,可更有效地摄取 养料,以满足其生长发育的需要。 图 2.3 霉菌菌丝 A. 无隔多核菌丝; B. 有隔单核菌丝; C. 有隔多核菌丝 1 、吸器 寄生真菌在寄主细胞间的菌丝常发生旁枝,侵入寄主细胞吸取养料,这种伸 入寄主细胞内的特殊菌丝分枝,称为吸器 ( 或吸胞 haustoria) 。吸器有各 种形状,如球状、指状、根状和丝状等 ( 图 2.4) 。一般专性寄生真菌,如 锈菌、霜霉菌、白粉菌等都有吸器。 图 2.4 真菌吸器的类型 A. 球状 B. 根状 C . 指状 D. 佛手状 2 、菌环和菌丝网 有些捕食性真菌,在菌丝分枝上形成环状菌丝,借以捕捉线虫,这叫菌 环。菌网是由菌丝形成的许多网眼所组成。每一网眼都极富于粘性,当线虫 与之接触,它就象捕蝇纸粘苍蝇一样,立刻把线虫粘住。然后从菌网的粘虫 处生出一小枝,穿透虫体而进入体内,吸收虫体内的营养物质 ( 图 2.5) 。 图 2.5 真菌的菌套和菌网 A. 菌套 B. 简单菌网 C. 复杂菌网 此外,还有些真菌的菌丝顶端常膨大甚至分枝,借以附着寄主或其它目 的物,称为附着枝。 匍匐丝和假根也是菌丝的适应性变态。 ( 四 ) 、菌丝的组织体 许多真菌在发育循环 ( 生活史 ) 的某 个阶段,菌丝常相互交织起来,组成一定的组 织体。常见的组织体有菌索、菌核和子座等。 1 、菌索 有些高等真菌的菌丝体平等排列组成长 条状似绳索,叫做“菌索”。菌索周围有外皮, 尖端是生长点,多生在树皮下或地下,根状, 白色或其它各种颜色。它有助于真菌迅速运送物质和蔓延侵染的功能,在不适宜的环境条件下呈休眠状态。 2 、菌核 菌核 (sclerotium) 是由菌丝体交织成团状的一种坚硬休眠体。它的外层由深色厚壁菌丝组成,内层由淡色菌丝 构成。不同真菌所产生的菌核,其形状和大小也各不相同,药用的茯苓、猪苓、茯神、雷丸和麦角等都是真菌的菌核。 引起水稻纹枯病的离心丝核菌 ( Rhizoctonia centrifuga ) 所形成的菌核小如油菜籽。大的茯苓重达 60kg 。在条 件适合时,菌核可萌发产生子实体,菌丝和分生孢子等 ( 图 2.5) 。 图 2.6 子座和菌核 3 、子座 子座 (stroma) 是由菌丝体组成的一种 垫座组织,有时是由真菌菌丝和寄主组织混 合构成。子座有垫状、壳状及其它各种形状。 在子座上面或里面产生各种子实体。子座和 子实体是连续形成的,故子座可称作繁殖体 的一部分 ( 图 2.6)。 (五) 、单细胞真菌 - 酵母菌 有一小部分真菌营养体外形不同于一般 霉菌,不形成菌丝,它们是圆形或卵圆形的 单细胞真菌,例如酵母菌。酵母菌无性繁殖 以出芽繁殖或分裂繁殖。出芽繁殖是酵母菌 最普遍的方式,先在细胞一端生一小突起, 叫生“芽”,当芽长到正常大小时,或脱离母细胞;或与母细胞相连接,在子细胞上又长出新芽,如此反复进行,最

后成为具有发达或不发达分枝状的假菌丝(见图2)。假菌丝与真菌丝不同,其两细胞间有一细腰,而不象真正菌 二、真的 院状又有酵母状细型形态 图2.7酵母菌的芽殖及菌丝 华阶段之后,讲入终殖阶段,经过婆产生许多新个体。直 的繁殖方式通常分为有性繁殖和无性繁殖二类。 有性繁殖以细胞核的结合为特? 无性繁殖是指不经过两性细胞的配合便能产生新的个体,即营养繁殖为特征。 分真菌都能进行无性与有性繁殖,并且以无性繁殖为主。有的菌种缺少无性繁殖阶 段,而另 一些菌种缺少有性繁殖阶段。 (一)、真菌的无性繁殖 1、无性繁殖的类型 真菌的无性繁殖方式可概括为四种:()菌丝体的断裂片段可以产生新个体,大多数真菌都能进行这种无性繁殖 实验室转管 接种便是利用这特月 就是这种类型 来殖菌种巴营养细分裂严为 生无性每个老发为新个体 一个新个体,酵母菌属的无性繁殖 、无性孢子的类型 无性繁殖过程所产生的孢子称无性孢子(见图2,8)。无性孢子的形状、颜色 细胞数目、排列方式、产生方法都有种的特征性,因而可作为鉴定菌种的依据。 图2.8真菌无性孢子的主要类群A.游动孢子B.孢囊孢子 A C.分生孢子D.芽孢子E.厚垣孢子F.节孢子 (1)游动孢子(zoospor©)鞭毛菌的菌丝可直接形成或发有成各种形状的游 0 动孢子囊,游动孢子囊内的原生质体分割成许多小块,小块逐渐变圆,围以薄膜 球形后部发形 根毛 在水中游动 个孢孢子,数量 0 (3)分生孢子(Conidio 成C m)子茵和半知菌类的无性徇子 是分生孢子。简单的分生孢子是由特化菌丝先端通过一定方式发生的,其下方的 特化菌丝叫分生孢子梗(conidio phore©)。梗有单生的,散生的,成丛的,还有成束的。有的分生孢子梗自垫状菌丝 生出,使产孢总体结构呈盘状,叫分生孢子盘。有的菌种是在菌丝堆上生出分生孢子梗和分生孢子。还有些真菌的分 生孢子生在覆碗状或球形的分生孢子器中。 ④)厚垣孢子(chlamydospore)有些真菌在菌丝顶端或中间产生一个孢子,它的外围被厚壁包围着,叫厚坦孢子 也叫厚 图2.9真菌的有性孢子 休眠孢子囊B。卵孢子C.接合孢子D.子囊孢子E.担孢子 ore) 有些真菌菌丝发生断裂后成为节孢子 、有性的商 @ 有性繁殖以细胞核的结合为特征,这种核的结合是通过能动或不能动的配子 有性繁殖过程 )质 (②)核配:两个细胞里的核进行配合 真菌从质配到核配之间时间有长有短,这段时间称双核期,即每个细胞里有两个没 A 有结合的核。这是真菌特有的现象。(③)减数分裂:核配后或迟或早将继之以减 数分裂,减数分裂使染色体数目减为单倍。真菌的有性生殖一般是通过性细胞的结 合,产生一定形态的有性孢子米实现的。真菌形成有性孢子有两种不同方式。第一 种方式是真菌经 0 减数 E 卵菌和接合菌的 D 然后年设性孢子:所以这种有性孢子 州屋核是处体阶段,子囊菌和担子菌的有性孢 2、有性孢子类型 (1)卵孢子(oospore)©卵菌中的有性孢子为卵孢子。当繁殖时在菌丝上先生出藏卵器和雄器,雄器的核移入藏 卵器与卵球结合后形成双倍体的卵孢子。在不同的菌种中卵球可能一个,也可能多个(图2.9) (2)接合孢子(zygospore)接合菌的有性孢子为接合孢子。来自两个不同菌株的同形配偶囊,互相接触后,接 触处的胞壁溶解,来自双方的细胞质和细胞核融合起来形成一个双倍体的接合孢子。 (3)子囊孢子(ascospor©)子囊菌的有性孢子为子囊孢子。双核菌丝产生幼小子囊,其中的双核进行核配后减 数分裂产生4个新核,再分裂一次形成8个核,然后以核为中心逐步形成单倍体的子囊孢子,一个子囊内往往有8个 子囊孢子
后成为具有发达或不发达分枝状的假菌丝 ( 见图 2.7) 。假菌丝与真菌丝不同,其两细胞间有一细腰,而不象真正菌 丝横隔处两细胞宽度一致。 图 2.7 酵母菌的芽殖及假菌丝 有些真菌既有菌丝状又有酵母状细胞形态。 二、真菌的繁殖体 真菌通过营养阶段之后,便进入繁殖阶段,经过繁殖产生许多新个体。真菌 的繁殖方式通常分为有性繁殖和无性繁殖二类。有性繁殖以细胞核的结合为特征, 无性繁殖是指不经过两性细胞的配合便能产生新的个体,即营养繁殖为特征。大部 分真菌都能进行无性与有性繁殖,并且以无性繁殖为主。有的菌种缺少无性繁殖阶 段,而另一些菌种缺少有性繁殖阶段。 (一) 、真菌的无性繁殖 1、无性繁殖的类型 真菌的无性繁殖方式可概括为四种:(1) 菌丝体的断裂片段可以产生新个体,大多数真菌都能进行这种无性繁殖, 实验室“转管”接种便是利用这一特点来繁殖菌种。(2) 营养细胞分裂产生子细胞,如裂殖酵母菌无性繁殖就象细菌 一样,母细胞一分为二的繁殖。(3) 出芽繁殖,母细胞出“芽”,每个“芽”成为一个新个体,酵母菌属的无性繁殖 就是这种类型的繁殖。(4) 产生无性孢子,每个孢子可萌发为新个体。 2、无性孢子的类型 无性繁殖过程所产生的孢子称无性孢子(见图 2.8) 。无性孢子的形状、颜色、 细胞数目、排列方式、产生方法都有种的特征性,因而可作为鉴定菌种的依据。 图 2.8 真菌无性孢子的主要类群 A. 游动孢子 B. 孢囊孢子 C. 分生孢子 D. 芽孢子 E. 厚垣孢子 F. 节孢子 (1) 游动孢子 (zoospore) 鞭毛菌的菌丝可直接形成或发育成各种形状的游 动孢子囊,游动孢子囊内的原生质体分割成许多小块,小块逐渐变圆,围以薄膜 而形成游动孢子。游动孢子肾形、梨形或球形,具一或二根鞭毛,在水中游动一 段时间后,鞭毛收缩,产生细胞壁进行休眠,然后萌发形成新个体。 (2) 孢囊孢子 (Sporangiospores) 接合菌无性繁殖所产生的孢子生在孢子 囊内,孢子囊一般生于营养菌丝或孢囊梗的顶端。孢子囊内的原生质体割裂成许 多小块,每一块发育成一个孢囊孢子,数量一般都相当大。 (3) 分生孢子 (Conidiospore 或 Conidium) 子囊菌和半知菌类的无性孢子 是分生孢子。简单的分生孢子是由特化菌丝先端通过一定方式发生的,其下方的 特化菌丝叫分生孢子梗 (conidio phore) 。梗有单生的,散生的,成丛的,还有成束的。有的分生孢子梗自垫状菌丝 生出,使产孢总体结构呈盘状,叫分生孢子盘。有的菌种是在菌丝堆上生出分生孢子梗和分生孢子。还有些真菌的分 生孢子生在覆碗状或球形的分生孢子器中。 (4) 厚垣孢子 (chlamydospore) 有些真菌在菌丝顶端或中间产生一个孢子,它的外围被厚壁包围着,叫厚垣孢子, 也叫厚壁孢子。 图 2.9 真菌的有性孢子 A. 休眠孢子囊 B. 卵孢子 C. 接合孢子 D. 子囊孢子 E. 担孢子 (5) 节孢子 (arthrospore) 有些真菌菌丝发生断裂后成为节孢子。 (二) 、真菌的有性繁殖 1、有性繁殖的过程 有性繁殖以细胞核的结合为特征,这种核的结合是通过能动或不能动的配子、 配偶囊、菌体之间的结合来实现的。有性繁殖过程一般包括下列三个阶段: (1) 质 配 :首先是两个细胞的原生质进行配合。 (2) 核配 :两个细胞里的核进行配合。 真菌从质配到核配之间时间有长有短,这段时间称双核期,即每个细胞里有两个没 有结合的核。这是真菌特有的现象。 (3) 减数分裂 :核配后或迟或早将继之以减 数分裂,减数分裂使染色体数目减为单倍。真菌的有性生殖一般是通过性细胞的结 合,产生一定形态的有性孢子来实现的。真菌形成有性孢子有两种不同方式。第一 种方式是真菌经过核配以后,含有双倍体细胞核的细胞直接发育而形成有性孢子, 这种孢子的细胞核处于双倍体阶段,在它萌发时才进行减数分裂,卵菌和接合菌的 有性孢子就是这种情况,处于双倍体阶段。第二种方式是在核配以后,双倍体的细 胞核进行减数分裂,然后再形成有性孢子,所以这种有性孢子的细胞核是处于单倍体阶段,子囊菌和担子菌的有性孢 子属于这种情况。 2、有性孢子类型 (1) 卵孢子 (oospore) 卵菌中的有性孢子为卵孢子。当繁殖时在菌丝上先生出藏卵器和雄器,雄器的核移入藏 卵器与卵球结合后形成双倍体的卵孢子。在不同的菌种中卵球可能一个,也可能多个 ( 图 2.9) 。 (2) 接合孢子 (zygospore) 接合菌的有性孢子为接合孢子。来自两个不同菌株的同形配偶囊,互相接触后,接 触处的胞壁溶解,来自双方的细胞质和细胞核融合起来形成一个双倍体的接合孢子。 (3) 子囊孢子 (ascospore) 子囊菌的有性孢子为子囊孢子。双核菌丝产生幼小子囊,其中的双核进行核配后减 数分裂产生 4 个新核,再分裂一次形成 8 个核,然后以核为中心逐步形成单倍体的子囊孢子,一个子囊内往往有 8 个 子囊孢子

(4)担孢子(6 asidiospore)担子菌的有性孢子为担孢子。担孢子的形成过程与子囊孢子相似,不同的是①核配后 减数分裂所形成的4个核不再进行分裂:②以核为中心所形成的担孢子最终在担子外部形成:③担子有纵隔的, 单室无隔: 工培养基上由一段 一丛)菌丝或一个(或一堆 后商的商落和放线记十你定 女 单组 丝较粗面生长很快 商解保去证的心米生结品保 落没有固定大小 有的霉菌菌落生长则有一定的局限性, 呈现不同颜色:一般处于菌落中心的南丝南龄较大,位于边缘的则较年幼。菌落具有“霉味”。同一种霉菌,在不同 成分培养基上的菌落特征可能有变化。但各种霉菌,在同一培养基上的菌落形状、颜色等相对稳定。故菌落特征也是 竖定霉茵的重要依据之一 酵母菌的菌落与细菌的有些相似,但较细菌菌落大而厚,一般呈油脂或蜡脂状,表面光滑、湿润、呈乳白色或红色。 有些种的菌落可因培养时间过长而表面皱缩。酵母菌茵落往往有“酒香味” 表2-3真菌分类系统比钱 第二的主类 八米 Ainsworth et al.(1973)真菌字典(1995)】 真菌的分类系统很多,各派分类论点各不相同,下面仅介绍 真菌界 其南界Fung1) 其中的两种较有代表性的真菌分类系统(表2-3 粘菌门(裸菌门) 壶菌门 1、安斯沃思(G.C.Ainsworth1971,1973)的分类系统 真菌门 接合菌门 该系结在Whittake将直茵独立成界的基出上,将直菌界 为两个门(真菌门和粘菌门),在真菌门内根据有性孢子的类 鞭毛南亚门 接合璃纲 型、菌丝是否有隔膜等性状分为5个亚门,即鞭毛菌亚门、接 纲 毛菌纲 合菌亚门、子囊菌亚门、担子菌亚门和半知菌亚门。这一分类系 子囊菌门 肿菌纲 担子菌门 分类 箱纲 担子菌纲 真菌字典》的分类系统 1995 据16sRNA序列的研究、生物化学和细胞壁 接合南亚门 冬孢菌纲 接合带铜 组分以及DNA序列分析的结果,国际真菌学研究的权威机构 黑粉菌纲 英国国际其南研究所(International Mycological 毛菌纲 藻界(Chromista】 Institute)出版的第8版《真菌字典》中,将原来的真菌界 子囊菌亚门 丝壶菌门 划分为原生动物界、藻界和真菌界。真南界仅包括了4个门, 半子囊茵纲 网粘菌门 即壶菌门、接合菌门、子囊菌门和担子菌门。卵菌、丝壶菌和网 不整囊菌纲 卵菌门 和褐藻类具有近缘关系 核菌纲 原生动物界(Protozoa】 腔菌纲 集胞菌门 自然分类”仍需探讨。 真菌的代表属 虫囊菌 网柄菌门 一)、卵菌 南纲 粘南门 腐暴属(Pythium 担子菌亚门 根肿菌门 腐霉菌丝体在培养基上或 冬孢菌纲 瓜果上集生,呈白绒毛状 很象棉花。在显微镜下无 层菌纲 色透明、无多核、有分 腹菌纲 半知菌亚门 芽孢纲 孰后 丝孢环 条件适合球形的 囊上很快生出 腔孢纲 泡囊,孢子囊里的内含物迅速流入泡囊,然后在泡囊内分化成游动孢子。游动孢 子常为肾形,侧面凹处生两根鞭毛,成熟时泡囊破裂,孢子四散(图2.10) 图2.10德巴利腐莓(Pythium debaryanum)的形态特征和发有循环 1、营养茵丝2、孢子囊、泡囊4、游动孢子5、游动孢子休止 6、萌发7、藏卵器和雄器8、质配9、核配10、卵孢子11、萌发
(4) 担孢子 (basidiospore) 担子菌的有性孢子为担孢子。担孢子的形成过程与子囊孢子相似,不同的是① 核配后 减数分裂所形成的 4 个核不再进行分裂; ② 以核为中心所形成的担孢子最终在担子外部形成;③ 担子有纵隔的, 也有生横隔的,多数是单室无隔。 三、真菌的菌落特征 在自然基质或人工培养基上由一段 ( 或一丛 ) 菌丝或一个 ( 或一堆 ) 孢子发展而成的菌丝体的整体称菌落。 霉菌的菌落和放线菌一样也是由分枝状菌丝组成。因菌丝较粗而长,形成的菌落较疏松,呈绒毛状、絮状或蜘蛛网状, 一般比细菌菌落大几倍到几十倍。有些霉菌,如根霉、毛霉、链孢霉生长很快,菌丝在固体培养基表面蔓延,以至菌 落没有固定大小 。有的霉菌菌落生长则有一定的局限性,直径 1~2cm 或更小。菌落表面常呈现出肉眼可见的不同结 构和色泽特征,这是因为霉菌形成的孢子有不同形状、构造和颜色,有的水溶性色素可分泌到培养基中,使菌落背面 呈现不同颜色;一般处于菌落中心的菌丝菌龄较大,位于边缘的则较年幼。菌落具有“霉味”。同一种霉菌,在不同 成分培养基上的菌落特征可能有变化。但各种霉菌,在同一培养基上的菌落形状、颜色等相对稳定。故菌落特征也是 鉴定霉菌的重要依据之一。 酵母菌的菌落与细菌的有些相似,但较细菌菌落大而厚,一般呈油脂或蜡脂状,表面光滑、湿润、呈乳白色或红色。 有些种的菌落可因培养时间过长而表面皱缩。酵母菌菌落往往有“酒香味”。 表 2-3 真菌分类系统比较 第二节 真菌的主要类群 一、真菌的分类系统 真菌的分类系统很多,各派分类论点各不相同,下面仅介绍 其中的两种较有代表性的真菌分类系统(表 2-3 )。 1 、安斯沃思 (G. C. Ainsworth 1971,1973) 的分类系统 该系统在 Whittake 将真菌独立成界的基础上,将真菌界分 为两个门 ( 真菌门和粘菌门 ) ,在真菌门内根据有性孢子的类 型、菌丝是否有隔膜等性状分为 5 个亚门,即鞭毛菌亚门、接 合菌亚门、子囊菌亚门、担子菌亚门和半知菌亚门。这一分类系 统在上个世纪较有影响,但显然这一系统仍属“人为分类”而非 真正按亲缘关系和客观反应系统发育关系对真菌进行的“自然 分类”。 2 、《真菌字典》的分类系统 1995 年 , 根据 16s RNA 序列的研究、生物化学和细胞壁 组分以及 DNA 序列分析的结果,国际真菌学研究的权威机构— 英国国际真菌研究所( International Mycological Institute )出版的第 8 版《真菌字典》中,将原来的真菌界 划分为原生动物界、藻界和真菌界。真菌界仅包括了 4 个门, 即壶菌门、接合菌门、子囊菌门和担子菌门。卵菌、丝壶菌和网 粘菌被发现与硅藻类和褐藻类具有近缘关系,这一类群被称为藻 界,而其它粘菌被认为属于原生动物界。《真菌字典》的分类系 统较之安斯沃思的分类系统有了进步,但是否是代表了真正的 “自然分类”仍需探讨。 二、真菌的代表属 (一)、卵菌 腐霉属 ( Pythium ) 腐霉菌丝体在培养基上或 瓜果上集生,呈白绒毛状, 很象棉花。在显微镜下无 色透明、无隔多核、有分 枝。孢子囊有管状或膨大 的管状及球状两类,没有 孢囊梗的分化,孢子囊成 熟后产生游动孢子或芽 管。在条件适合时,孢子 囊上很快生出一个球形的 泡囊,孢子囊里的内含物迅速流入泡囊,然后在泡囊内分化成游动孢子。游动孢 子常为肾形,侧面凹处生两根鞭毛,成熟时泡囊破裂,孢子四散 (图 2.10) 。 图 2.10 德巴利腐霉 ( Pythium debaryanum ) 的形态特征和发育循环 1 、营养菌丝 2 、孢子囊 、泡囊 4 、游动孢子 5 、游动孢子休止 6 、萌发 7 、藏卵器和雄器 8 、质配 9 、核配 10 、卵孢子 11 、萌发 Ainsworth et al .(1973) 真菌字典( 1995 ) 真菌界 真菌界 (Fungi) 粘菌门(裸菌门) 壶菌门 真菌门 接合菌门 鞭毛菌亚门 接合菌纲 菌纲 毛菌纲 壶菌纲 子囊菌门 肿菌纲 担子菌门 菌纲 担子菌纲 接合菌亚门 冬孢菌纲 接合菌纲 黑粉菌纲 毛菌纲 藻界(Chromista) 子囊菌亚门 丝壶菌门 半子囊菌纲 网粘菌门 不整囊菌纲 卵菌门 核菌纲 原生动物界(Protozoa) 腔菌纲 集胞菌门 虫囊菌纲 网柄菌门 盘菌纲 粘菌门 担子菌亚门 根肿菌门 冬孢菌纲 层菌纲 腹菌纲 半知菌亚门 芽孢纲 丝孢纲 腔孢纲

腐有性生殖产生液卵器和雄器。卵器分化为卵球和卵周质,藏卵器原来是多核的,在分化时除留一核于卵球 转移到 核外 器内西的 ,外 其余的逐新解体。金合时器的细胞核和细 管 牙 生孢子 有的府还能转换省庆化合物 孢囊梗和幼年孢子囊2 子囊破裂后露出囊轴和孢囊孢子3 接合蘭 图211高大毛莓 1。手层属(场0r 毛蛋的菌丝体呈棉寥状,在基物上或基物内能广污莓证。菌丝无隔名核,幼嫩 时,原生质浓稠、均匀一致,老时则出现液泡并含有各种内含物。 毛霉的无性繁殖:在毛霉的培养中,可见到气生菌丝加长,先端膨大,成为具 有一个头部的长丝: (sporangium) 满很多细胞移 面的陶 叫孢 有 11 孢子成熟 来。孢囊孢 在空气中被吹散 遇到适宜环境,萌发而形成新的菌丝体 称为原配子壶 原配子壶接钟后。顶端各白影大并形成描阻。隔成一细胞。此细胞叫图子查(。ametang》。相接轴的两司子素之信 的横隔消失,经过质配、核配,同时外部形成厚壁,即形成接合孢子。接合孢子孢壁很厚,褐色,表面粗糙有突出物! 接合孢子经一段休眠期后才能萌发。萌发前减数分裂,萌发时,孢壁破裂,长出芽管,芽管顶端形成一孢子囊,在孢 子囊内产生大量单倍体的孢囊孢子(图2.12)。图2.12毛霉属的生活史 三霉的有性结合可分为异宗配合或同宗配合。凡是由不同性菌丝体上形成的性 器官结合而形成 孢子则称为同宗配合。 般用“由同 和 的生活史如下 丝一〉孢子囊一〉孢囊孢子一〉菌丝 有性繁 (+)菌丝-〉(+)配子囊 ()菌丝一)配子囊 接合孢子一发芽→孢子囊一孢囊孢子一南丝 毛蛋在自织界分 广 ,土壤、空气中都有很多毛霉孢子。多种毛霉能产生蛋 白酵,我固多用来做豆腐气:的显是用总状毛 sus)制作的 )产生乳酸、琥珀酸及甘油 毛霉还能对甾族化合物起转化作用。有些毛霉能引起谷物、果品、蔬菜和其他食品 的腐败。 图213根萄 (rhizoid和 ,很多特 生长 由营养茵丝体 立生形的 生出假根与基物 接蚀」 假根相对处向上 个从生于担上(图213) 色。孢囊孢子球形、卵形或不规则,或有棱角或有线纹、无色或浅褐色、蓝灰 鱼等。接合孢子由菌丝体或细铜阁南丝生出配子童,由两个同形对生的配子囊结 合而成。根霉属的菌除有性根霉(R.sexualis)为同宗配合外,已知的其他 种都是异宗配合(图2.14)。 根霉在自然界分布很广,空气、土壤以及各种器皿表面都有存在。并常出 围包 甘薯等 力相当强 化南,同时也是家 (=) 子 子垂菌的大多数 种类形成菌丝,菌丝右隔膜。子囊茵的无性繁殖主要是产生分生狗子而酵母菌及其他少数子 囊菌则以芽殖、裂殖为正常繁殖形式。有性繁殖产生子囊孢子,子囊孢子生于子囊(as©uS)内。子囊是一种囊状结构, 球形、棒形或圆筒形,因种而异。典型的子囊内有8个子囊孢子。子囊有4种若生方式(图2.15),(a)有性 酵母菌的子囊往往为裸露的。(b)者生在闭囊壳(peritheciuon)内,闭囊壳为完全封闭式,呈球形。(c)着 生在子囊壳(perithecium)内,子囊壳似烧瓶形,有孔口。(d)着生在子囊盘(apothecium)中,子囊盘为开 口的、盘状。闭囊壳、子囊壳、子囊盘统称为子囊果(asco心rD)。丝状真南的子囊一般被包被于子囊果内
腐霉有性生殖产生藏卵器和雄器。藏卵器分化为卵球和卵周质,藏卵器原来是多核的,在分化时除留一核于卵球 内,其余的均转移到卵周质层逐渐分解。雄器最初也是多核的,除一核外其余的逐渐解体。配合时雄器的细胞核和细 胞质经由授精管转入藏卵器内,两核结合形成卵孢子,外表光滑或有刺。卵孢子萌发通常生芽管,在芽管顶端生孢子囊。 腐霉有水生的、两栖的、陆生的、腐生和兼寄生的各种类型。腐霉能合成生物素、叶酸、泛酸、核黄素、抗坏血 酸等维生素;腐霉能分泌果胶酶、纤维素酶等多种酶类;有的腐霉还能转换甾族化合物;一些种类引起幼苗猝倒病, 瓜果绵腐病等。 1 、孢囊梗和幼年孢子囊 2 、孢子囊破裂后露出囊轴和孢囊孢子 3 、接合孢子 (二)、接合菌 图 2.11 高大毛霉 1 、毛霉属 ( Mucor ) 毛霉的菌丝体呈棉絮状,在基物上或基物内能广泛蔓延。菌丝无隔多核,幼嫩 时,原生质浓稠、均匀一致,老时则出现液泡并含有各种内含物。 毛霉的无性繁殖:在毛霉的培养中,可见到气生菌丝加长,先端膨大,成为具 有一个头部的长丝,头部下生一隔膜,将头部与长丝分开。头部发育为孢子囊 (sporangium) ,囊内充满很多细胞核,每个细胞核周围细胞质浓缩,外面形成孢壁, 就成为孢囊孢子。孢子囊下面的菌丝叫孢囊梗 (sporangiophore) ;孢囊梗突入孢 子囊内的部分,叫做囊轴(columella)。孢囊梗和囊轴间还有囊托(apophysis)(见图 2.11) 。孢子囊成熟,囊壁破裂,孢囊孢子分散出来。孢囊孢子在空气中被吹散, 遇到适宜环境,萌发而形成新的菌丝体。毛霉的有性繁殖:相邻近的两菌丝各自向对方生出极短侧枝,称为原配子囊, 原配子囊接触后,顶端各自膨大并形成横隔,隔成一细胞,此细胞叫配子囊 (gametangium) 。相接触的两配子囊之间 的横隔消失,经过质配、核配,同时外部形成厚壁,即形成接合孢子。接合孢子孢壁很厚,褐色,表面粗糙有突出物。 接合孢子经一段休眠期后才能萌发。萌发前减数分裂,萌发时,孢壁破裂,长出芽管,芽管顶端形成一孢子囊,在孢 子囊内产生大量单倍体的孢囊孢子 ( 图 2.12) 。图 2.12 毛霉属的生活史 毛霉的有性结合可分为异宗配合或同宗配合。凡是由不同性菌丝体上形成的性 器官结合而形成有性孢子,称为异宗配合;由同一个菌丝体上形成的配子囊结合而 产生有性孢子则称为同宗配合。一般用“十”号和“一”号分别代表两个性别不同 的菌丝体。 毛霉的生活史如下: 无性繁殖:菌丝--> 孢子囊--> 孢囊孢子--> 菌丝 有性繁殖: (+) 菌丝 --> (+)配子囊 接合孢子 →发芽→孢子囊→孢囊孢子→菌丝 (-) 菌丝 --> (-) 配子囊 毛霉在自然界分布广泛,土壤、空气中都有很多毛霉孢子。多种毛霉能产生蛋 白酶,我国多用来做豆腐乳。四川的豆豉就是用总状毛霉 ( M.racemosus ) 制作的。 许多毛霉产生有机酸,如鲁氏毛霉 ( M.rouxianus ) 产生乳酸、琥珀酸及甘油等。 毛霉还能对甾族化合物起转化作用。有些毛霉能引起谷物、果品、蔬菜和其他食品 的腐败。 2 、根霉属 ( Rhizopus ) 图 2.13 根霉 根霉与毛霉同属毛霉目 (Mucorales) ,很多特征相似。主要区别在于根霉有 假根 (rhizoid) 和匍匐菌丝 (stolon) 。根霉在培养基或自然基物上生长时, 由营养菌丝体产生弧形的匍匐菌丝向四周蔓延。并由匍匐菌丝生出假根与基物 接触。与假根相对处向上生长出孢囊梗,顶端形成孢子囊,内生孢囊孢子。孢 囊梗不分枝,直立,二、三个丛生于假根上 ( 图 2.13) 。孢子囊成熟时呈黑 色。孢囊孢子球形、卵形或不规则,或有棱角或有线纹、无色或浅褐色、蓝灰 色等。接合孢子由菌丝体或匍匐菌丝生出配子囊,由两个同形对生的配子囊结 合而成。根霉属的菌除有性根霉 ( R.sexualis ) 为同宗配合外,已知的其他 种都是异宗配合 ( 图 2.14) 。 根霉在自然界分布很广,空气、土壤以及各种器皿表面都有存在。并常出 现于淀粉食品上,引起馒头、面包、甘薯等发霉变质,或造成水果蔬菜腐烂。 根霉的用途很广,在我国用它们制曲酿酒已有悠久历史。有的根霉的淀粉酶活力相当强,多用作糖化菌,同时也是家 用甜酒曲的主要菌种。近年来在甾体激素转化、有机酸 ( 延胡索酸 、乳酸 ) 的生产中被广泛利用。 ( 三 ) 、子囊菌 子囊菌的大多数种类形成菌丝,菌丝有横隔膜。子囊菌的无性繁殖主要是产生分生孢子,而酵母菌及其他少数子 囊菌则以芽殖、裂殖为正常繁殖形式。有性繁殖产生子囊孢子,子囊孢子生于子囊 (ascus) 内。子囊是一种囊状结构, 球形、棒形或圆筒形,因种而异。典型的子囊内有 8 个子囊孢子。子囊有 4 种着生方式 ( 图 2.15) ,( a )有性 酵母菌的子囊往往为裸露的。( b )着生在闭囊壳 (peritheciuon) 内,闭囊壳为完全封闭式,呈球形。( c )着 生在子囊壳( perithecium )内,子囊壳似烧瓶形,有孔口。( d )着生在子囊盘 (apothecium) 中,子囊盘为开 口的、盘状。闭囊壳、子囊壳、子囊盘统称为子囊果 (ascocarp) 。丝状真菌的子囊一般被包被于子囊果内