
酶和过氧化物酶)的活化剂,尤其是影响糖酵解和三羧酸循环。锰使光合作用中 水裂解为氧。缺锰时,叶脉间缺绿,伴随小坏死点的产生。缺绿会在嫩叶或老叶 出现,以植物种类和生长速率而定。 1I、钠钠离子(Na)在C4和CAM植物中催化PEP的再生。钠离子对许 多C3植物的生长也是有益的,它使细胞膨胀从而促进生长。钠还可以部分地代 替钾的作用,提高细胞液的渗透势。缺钠时,这些植物呈现黄化和坏死现象,甚 至不能开花。 (四)第4组一一参与氧化还原反应的营养 12、铁铁是光合作用、生物固氨和呼吸作用中的细胞色素和非血红素铁蛋 白的组成。铁在这些代谢方面的氧化还原过程中都起赠子传递作用。由于叶绿 体的某些叶绿素一蛋白复合体合成需要铁,所以,缺铁时会出现叶片叶脉间缺绿。 与缺镁症状相反,缺铁发生于嫩叶,因铁不易从老叶转移出来,缺铁过甚或过久 时,叶脉也缺绿,全叶白化,华北果树的“黄叶病”就是植株缺铁所致。 13、锌锌离子(Z2+)是乙醇脱氢酶、谷氨酸脱氢酶和碳酸酐酶等的组成之 一。缺锌植物失去合成色氨酸的能力,而色氨酸是吲哚乙酸的前身,因此缺锌植 物的吲哚乙酸含量低。锌是叶绿素生物合成的必需元素。锌不足时,植株茎部节 间短,莲丛状,叶小且变形,叶缺绿。吉林和云南等省玉米“花白叶病”,华北 地区果树“小叶病”等都是缺锌的缘故。 14、铜铜是某些氧化酶(例如抗坏血酸氧化酶、酪氨酸酶等)的成分,可 以影响氧化还原过程(Cu*←一Cu+)。铜又存在于叶绿体的质体蓝素 中,后者是光合作用电子传递体系的一员。缺铜时,叶黑绿,其中有坏死点,先 从嫩叶叶尖起,后沿叶缘扩展到叶基部,叶也会卷皱或畸形。缺铜过甚时,叶脱 落。 15、镍镍是脲酶的金属成分,脲酶的作用是催化尿素水解成CO2和NH*。 镍也是氢化酶的成分之一,它在生物固氮中产生氢气起作用。缺镍时,叶尖积累 较多的脲,出现坏死现象。 16、钼钼离子(Mo+Mo6+)是硝酸还原酶的金属成分,起着电子传递作 用。钼又是固氮酶中钼铁蛋白的成分,在固氮过程中起作用。所以,钼的生理功 能突出表现在氮代谢方面。钼对花生、大豆等豆科植物的增产作用显著。缺钼时, 31
31 酶和过氧化物酶)的活化剂,尤其是影响糖酵解和三羧酸循环。锰使光合作用中 水裂解为氧。缺锰时,叶脉间缺绿,伴随小坏死点的产生。缺绿会在嫩叶或老叶 出现,以植物种类和生长速率而定。 11、钠 钠离子(Na+)在 C4 和 CAM 植物中催化 PEP 的再生。钠离子对许 多 C3 植物的生长也是有益的,它使细胞膨胀从而促进生长。钠还可以部分地代 替钾的作用,提高细胞液的渗透势。缺钠时,这些植物呈现黄化和坏死现象,甚 至不能开花。 (四)第 4 组——参与氧化还原反应的营养 12、铁 铁是光合作用、生物固氮和呼吸作用中的细胞色素和非血红素铁蛋 白的组成。铁在这些代谢方面的氧化还原过程中都起着电子传递作用。由于叶绿 体的某些叶绿素—蛋白复合体合成需要铁,所以,缺铁时会出现叶片叶脉间缺绿。 与缺镁症状相反,缺铁发生于嫩叶,因铁不易从老叶转移出来,缺铁过甚或过久 时,叶脉也缺绿,全叶白化,华北果树的“黄叶病”就是植株缺铁所致。 13、锌 锌离子(Zn 2+)是乙醇脱氢酶、谷氨酸脱氢酶和碳酸酐酶等的组成之 一。缺锌植物失去合成色氨酸的能力,而色氨酸是吲哚乙酸的前身,因此缺锌植 物的吲哚乙酸含量低。锌是叶绿素生物合成的必需元素。锌不足时,植株茎部节 间短,莲丛状,叶小且变形,叶缺绿。吉林和云南等省玉米“花白叶病”,华北 地区果树“小叶病”等都是缺锌的缘故。 14、铜 铜是某些氧化酶(例如抗坏血酸氧化酶、酪氨酸酶等)的成分,可 以 影 响 氧 化 还 原 过 程 ( Cu+ Cu 2+)。铜又存在于叶绿体的质体蓝素 中,后者是光合作用电子传递体系的一员。缺铜时,叶黑绿,其中有坏死点,先 从嫩叶叶尖起,后沿叶缘扩展到叶基部,叶也会卷皱或畸形。缺铜过甚时,叶脱 落。 15、镍 镍是脲酶的金属成分,脲酶的作用是催化尿素水解成 CO2 和 NH4 +。 镍也是氢化酶的成分之一,它在生物固氮中产生氢气起作用。缺镍时,叶尖积累 较多的脲,出现坏死现象。 16、钼 钼离子(Mo4+~ Mo6+)是硝酸还原酶的金属成分,起着电子传递作 用。钼又是固氮酶中钼铁蛋白的成分,在固氮过程中起作用。所以,钼的生理功 能突出表现在氮代谢方面。钼对花生、大豆等豆科植物的增产作用显著。缺钼时, +e Fe3+ Fe2+ -e

老叶叶脉间缺绿,坏死。而在花椰菜,叶皱卷甚至死亡,不开花或花早落。 三、作物缺乏矿质元素的诊断 (一)病症诊断法 植物缺乏矿质元素的病征检索表 病征 缺乏元素 A、老叶病征 B、病征常遍布整株,基部叶片干焦和死亡 C、植物浅绿,基部叶片黄色,干燥时呈褐色,茎部而细. . C、植株深绿,常呈红或紫色,基部叶片黄色,干燥时暗绿,茎短而细. B、病征常限于局部,基部叶片不干焦但杂色或缺绿,叶缘杯状卷起或卷皱 C、叶杂色或缺绿,有时呈红色,有坏死斑点,茎细. C、叶杂色或缺绿,在叶脉间或叶尖和叶缘有坏死小斑点,茎细.钾 C、坏死斑点大而普遍出现于叶脉间,最后出现于叶脉,叶厚,茎短.锌 A、嫩叶病征 B、顶芽死亡,嫩叶变形和坏死 C、嫩叶初呈钩状,后从叶尖和叶缘向内死亡. .钙 C、嫩叶基部浅绿,从叶基起枯死,叶捻曲. B、顶芽仍活但缺绿或萎蔫,无坏死斑点 C、嫩叶菱焉,无失绿,茎尖弱. C、嫩叶不菱蔫,有失绿 D、坏死斑点小,叶脉仍绿 D、无坏死斑点 E、叶耿仍绿.铁 E、叶脉失绿. .疏 (二)化学分析诊断法 化学分析是营养诊断的另一种根据。 第二节植物细胞对矿质元素的吸收(Asorption of mineral elements
32 老叶叶脉间缺绿,坏死。而在花椰菜,叶皱卷甚至死亡,不开花或花早落。 三、作物缺乏矿质元素的诊断 (一)病症诊断法 植物缺乏矿质元素的病征检索表 病 征 缺 乏 元 素 A、老叶病征 B、病征常遍布整株,基部叶片干焦和死亡 C、植物浅绿,基部叶片黄色,干燥时呈褐色,茎部而细.氮 C、植株深绿,常呈红或紫色,基部叶片黄色,干燥时暗绿,茎短而细. 磷 B、病征常限于局部,基部叶片不干焦但杂色或缺绿,叶缘杯状卷起或卷皱 C、叶杂色或缺绿,有时呈红色,有坏死斑点,茎细.镁 C、叶杂色或缺绿,在叶脉间或叶尖和叶缘有坏死小斑点,茎细.钾 C、坏死斑点大而普遍出现于叶脉间,最后出现于叶脉,叶厚,茎短.锌 A、嫩叶病征 B、顶芽死亡,嫩叶变形和坏死 C、嫩叶初呈钩状,后从叶尖和叶缘向内死亡.钙 C、嫩叶基部浅绿,从叶基起枯死,叶捻曲.硼 B、顶芽仍活但缺绿或萎蔫,无坏死斑点 C、嫩叶萎蔫,无失绿,茎尖弱.铜 C、嫩叶不萎蔫,有失绿 D、坏死斑点小,叶脉仍绿.锰 D、无坏死斑点 E、叶脉仍绿.铁 E、叶脉失绿.硫 (二)化学分析诊断法 化学分析是营养诊断的另一种根据。 第二节 植物细胞对矿质元素的吸收(Asorption of mineral elements

by plant cells) 细胞除了吸收水分外,还要从环境中吸收养料,借示踪原子法研究得知,不 仅无机物质的离子能进入细胞,分子量较大的有机物(如氨基酸、维生素等)也 能进入细胞。植物细胞与外界环境进行的一切物质交换,都必须通过各种生物膜, 特别是质膜。 一、生物膜(biological membrane) 植物细胞的原生质体是被质膜包围着,在细胞质和液泡之间,又有液泡膜隔 开。植物细胞里有许多细胞器,它们都是有膜包围着或者是由膜组成的。因此, 从某种意义上说,植物细胞是一个由膜系统组成的单位,这些膜把各种细胞器与 其他部分分隔开,有利于各细胞器分别行使各自特有的功能,有利于有秩序地、 有条不紊地进行各种代谢活动。研究表明,许多酶是埋藏在膜里或与膜结合在一 起,细胞许多生理、生化活动是在膜上或邻近的空间进行的。所以可以说,膜是 植物生理活动的中心所在。 (一)膜的特性和化学成分(compositions) 人们早期研究各种物质通过质膜的特性,发现细胞质膜具有让物质通过的性 质,称为透性(permeability)。但是质膜对各种物质的通过难易不一,有些容易 通过,有些则不易或不能通过,所以质膜对各种物质具有选择透性(selective permeability)。研究表明,膜对水的透性最大,水可以自由通过:越容易溶解于 脂质的物质,透性越大。所以膜一定是由亲水性物质和脂类物质组成。 膜的基本成分是蛋白质、脂质和糖。蛋白质约占30%~40%,脂质约占40%~ 60%,糖类10%~20%。膜内蛋白质是糖蛋白、脂蛋白等,它们起者结构、运输 及传递信息等方面的作用。脂质的主要成分是磷脂,包括磷脂酰胆碱、磷脂酰乙 醇胺,磷脂酰甘油和磷脂酰肌醇。磷脂的结构和形状如图21。磷脂既有两条易 溶于脂肪性溶剂中的非极性疏水“长尾巴”(通常为16~18个碳原子的脂肪酸侧 链),又有一个易溶于水的极性头部,所以磷脂是双亲媒性的化合物。磷脂是各 种膜的骨架,可能有调控细胞多种功能的作用。类囊体膜中还含有大量糖脂,主 要是半乳糖甘油二脂和双半乳糖甘油二脂。此外,膜上还含有固醇,夹杂在磷脂 之内
33 by plant cells) 细胞除了吸收水分外,还要从环境中吸收养料,借示踪原子法研究得知,不 仅无机物质的离子能进入细胞,分子量较大的有机物(如氨基酸、维生素等)也 能进入细胞。植物细胞与外界环境进行的一切物质交换,都必须通过各种生物膜, 特别是质膜。 一、生物膜(biological membrane) 植物细胞的原生质体是被质膜包围着,在细胞质和液泡之间,又有液泡膜隔 开。植物细胞里有许多细胞器,它们都是有膜包围着或者是由膜组成的。因此, 从某种意义上说,植物细胞是一个由膜系统组成的单位,这些膜把各种细胞器与 其他部分分隔开,有利于各细胞器分别行使各自特有的功能,有利于有秩序地、 有条不紊地进行各种代谢活动。研究表明,许多酶是埋藏在膜里或与膜结合在一 起,细胞许多生理、生化活动是在膜上或邻近的空间进行的。所以可以说,膜是 植物生理活动的中心所在。 (一) 膜的特性和化学成分(characteristics and compositions) 人们早期研究各种物质通过质膜的特性,发现细胞质膜具有让物质通过的性 质,称为透性(permeability)。但是质膜对各种物质的通过难易不一,有些容易 通过,有些则不易或不能通过,所以质膜对各种物质具有选择透性(selective permeability)。研究表明,膜对水的透性最大,水可以自由通过;越容易溶解于 脂质的物质,透性越大。所以膜一定是由亲水性物质和脂类物质组成。 膜的基本成分是蛋白质、脂质和糖。蛋白质约占 30%~40%,脂质约占 40%~ 60%,糖类 10%~20%。膜内蛋白质是糖蛋白、脂蛋白等,它们起着结构、运输 及传递信息等方面的作用。脂质的主要成分是磷脂,包括磷脂酰胆碱、磷脂酰乙 醇胺,磷脂酰甘油和磷脂酰肌醇。磷脂的结构和形状如图 2-1。磷脂既有两条易 溶于脂肪性溶剂中的非极性疏水“长尾巴”(通常为 16~18 个碳原子的脂肪酸侧 链),又有一个易溶于水的极性头部,所以磷脂是双亲媒性的化合物。磷脂是各 种膜的骨架,可能有调控细胞多种功能的作用。类囊体膜中还含有大量糖脂,主 要是半乳糖甘油二脂和双半乳糖甘油二脂。此外,膜上还含有固醇,夹杂在磷脂 之内
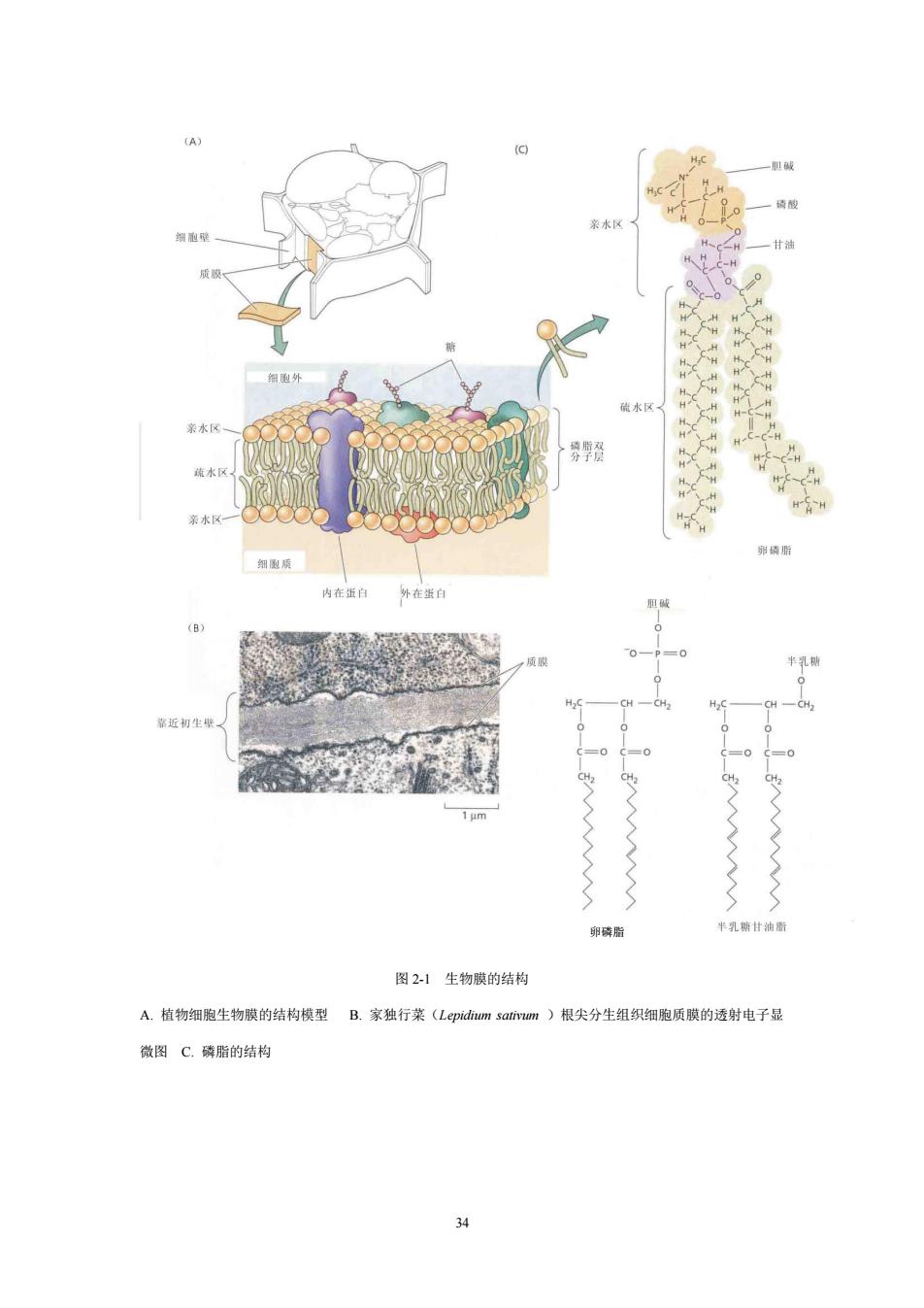
水 硫水 =0 卵磷脂 图21生物膜的结构 A.植物细胞生物膜的结构模型B.家独行菜(Lepidim sativum)根尖分生组织细胞质膜的透射电子显 微图C.磷脂的结构
34 图 2-1 生物膜的结构 A. 植物细胞生物膜的结构模型 B. 家独行菜(Lepidium sativum )根尖分生组织细胞质膜的透射电子显 微图 C. 磷脂的结构 卵磷脂

(二)膜的结构((structure of membrane) 生物膜的功能如此复杂,那么它的结构是什么样的呢?人们通过膜能流动、 断裂、重新组合、出芽或形成大泡囊后独立开来等现象,认识到膜是流动的,不 是静止的,它是不断适应细胞的生长活动而组成或发展的。膜之所以流动,与磷 脂分子的相对运动有关。膜在较高温度下,呈液相状态;在低温下即转变为固相 状态。自从1972年S.Singer和G.Nicolson提出流动镶嵌模型(fluid mosaic model) 以来,多年来人们从多方面的研究得以证实,生物膜结构的基本特点是:膜一般 是由磷脂双分子层(phospholipid bilayer)和镶嵌的蛋白质组成。磷脂分子的亲 水性头部位于膜的表面,疏水性尾部在膜的内部。膜上的蛋白质有些是与膜的外 表面相连,称为外在蛋白(extrinsic protein),亦称周围蛋白(peripheral protein): 有些是镶嵌在磷脂之间,甚至穿透膜的内外表面,称为内在蛋白(intrinsic protein),亦称整合蛋白(integral protein)(图2-1)。由于蛋白质在膜上的分布不 均匀,膜的结构不对称,部分蛋白质与多糖相连。膜脂和膜蛋白是可以运动的。 膜厚7~10nm。 二、细胞吸收溶质的方式和机理(Types and mechanisms) 根据目前的资料,植物细胞吸收溶质的方式共有4种类型:通道运输,载体 运输,泵运输和胞饮作用。 (一)通道运输(channel transport) 通道运输(channel transport)理论认为,细胞质膜上有内在蛋白构成的通道, 横跨膜的两侧。通道大小和孔内电荷密度等使得通道对离子运输有选择性,即一 种通道只允许某一种离子通过。通道蛋白有所谓“闸门”的结构,它的开和关决 定于外界信号。膜片钳技术(patch clamp technique)研究提示,某一种离子(例 如K+)在膜上有各种不同的通道,这些通道是否打开决定于不同电压范围或不 同的信号(如K+或Ca+浓度,pH值、蛋白激酶和磷酸酶等)。例如,当细胞外 的某一离子浓度比细胞内的该离子浓度高时,质膜上的离子通道被激活,通道闸 门打开,离子将顺着跨质膜的电化学势梯度进入细胞内(图2-2)。质膜上的通道 运输是一种简单扩散的方式,是一种被动运输(passive transport)
35 (二) 膜的结构(structure of membrane) 生物膜的功能如此复杂,那么它的结构是什么样的呢?人们通过膜能流动、 断裂、重新组合、出芽或形成大泡囊后独立开来等现象,认识到膜是流动的,不 是静止的,它是不断适应细胞的生长活动而组成或发展的。膜之所以流动,与磷 脂分子的相对运动有关。膜在较高温度下,呈液相状态;在低温下即转变为固相 状态。自从 1972 年 S.Singer 和 G. Nicolson 提出流动镶嵌模型(fluid mosaic model) 以来,多年来人们从多方面的研究得以证实,生物膜结构的基本特点是:膜一般 是由磷脂双分子层(phospholipid bilayer)和镶嵌的蛋白质组成。磷脂分子的亲 水性头部位于膜的表面,疏水性尾部在膜的内部。膜上的蛋白质有些是与膜的外 表面相连,称为外在蛋白(extrinsic protein),亦称周围蛋白(peripheral protein); 有些是镶嵌在磷脂之间,甚至穿透膜的内外表面,称为内在蛋白(intrinsic protein),亦称整合蛋白(integral protein)(图 2-1)。由于蛋白质在膜上的分布不 均匀,膜的结构不对称,部分蛋白质与多糖相连。膜脂和膜蛋白是可以运动的。 膜厚 7~10nm。 二、细胞吸收溶质的方式和机理(Types and mechanisms) 根据目前的资料,植物细胞吸收溶质的方式共有 4 种类型:通道运输,载体 运输,泵运输和胞饮作用。 (一)通道运输(channel transport) 通道运输(channel transport)理论认为,细胞质膜上有内在蛋白构成的通道, 横跨膜的两侧。通道大小和孔内电荷密度等使得通道对离子运输有选择性,即一 种通道只允许某一种离子通过。通道蛋白有所谓“闸门”的结构,它的开和关决 定于外界信号。膜片钳技术(patch clamp technique)研究提示,某一种离子(例 如 K+)在膜上有各种不同的通道,这些通道是否打开决定于不同电压范围或不 同的信号(如 K+或 Ca 2+浓度,pH 值、蛋白激酶和磷酸酶等)。例如,当细胞外 的某一离子浓度比细胞内的该离子浓度高时,质膜上的离子通道被激活,通道闸 门打开,离子将顺着跨质膜的电化学势梯度进入细胞内(图 2-2)。质膜上的通道 运输是一种简单扩散的方式,是一种被动运输(passive transport)