
酪乳杆菌)等细菌中含有,后者在Sapy e和(芽孢杆菌属)等细中含有。 甘袖磷壁酸的结构及其与肽聚塘分子中的胞壁酸的连接方式见图27。 P-0 甘四 世A @©囚 0K0 0 NH-CO-CH," H-co-CH, CH,tHC0-肽尾 图?甘油藻整酸的结构模式(东)技其伞体(雀线范晰内)的分f结构阁(有) ()甘油瞬酸套复单位 OH CH-0 OK 又=州婆部想 (2)葡糖酰甘油醉酸重夏单位: (③)N-乙酰葡葡胺-谛酸一甘油醉酸重复单位, CH.OR NHAC ON ()1,5-聚核糖醇确酸: °2 R=甘油酸 OH H-C-OR CH:-0 jt-30-35 2三甘碎壁酸和一种糖壁酸的结构

以下再将其余三种甘油磷壁酸和一种核醇磔壁酸的结构归纳在图2-8中。 障罐酸的主要生理功能有四:①因带负电荷,故可与环境中的Mg+等阳离子结合,提高 这些离子的浓度,以保证细胞膜上一些合戒瘦维持高活性的需要:②保证革兰氏阳性致病菌 (如A族链球菌)与其宿主间的粘连(主要为膜磷壁酸):③赋于革兰氏阳性以特异的表面 抗原:④提供某些噬菌体以特异的吸附受体。 革兰氏阴性细菌特有的脂多蟠(LPS,Hpopolysaccharide)结构脂多糖是位于革兰氏阴性 细菌细胞壁最外层的一层较厚(8~10mm)的类脂多糖类物质。它由类脂A、核心多糖和O特 异侧链三部分所组成。其主要功能有四:①是革兰氏阴性细菌致病物质一内毒素的物质基 韩:②与陵壁酸相似,也,有吸附Mg+、C+等阳离子以提高议些离子在细胞表面的浓度的作 用:③由于LS结构的变化,决定了革兰氏阴性细细胞表面抗原决定熊的多样性,例如,据 统计(1983),国际上已报道过的根据LPS的结构特性而鉴定过Saone(沙门氏菌稀)的表 面抗原类型多达2107个:④是许多噬菌体在细胞表面的吸附受体 脂多糖要维持其结构的稳定性需要足量Ca+的存在。如果用合剂去除Ca+,山PS就解 体。这时,革兰氏阴性细菌的内壁层肽聚糖就暴露出来,因而就可被溶菌酶所水解。 脂多糖分子的结构较复杂,现表解如下: (类脂A:2个N-乙酰葡糖蔽及5个长链脂肪酸 低心多起作e区6+8装 外楼心区:5个己糖(Hex),包括葡德胺、半乳糖、葡萄第 类脂A的结构在不同细菌中有所不同,大约有?一8种之多、它是革兰氏阴性细菌内毒素 CHOR -0 0- RO OR -0 OH 6 0特异侧链丁装心多精H0H (CHa) CH co R-( w或( :或 CH CH, CH, 91 0c0 (月佳酸) (棕钢酸) CH) CH, (豆蔻酸) 图?gLPS中类指A的分子结构(沙!幽属)

的毒性中心,其结构见图2-9。 在LPS的核心多糖区和O特异侧链区中有三种独特的糖,即2附3-脱氧辛糖酸(KDO) L甘油-D-甘露庚糖(Hep)和阿比可糖(Aba,即3,6-二脱氧D半乳糖),它们的结构见图2 10 CH.OR CH.OH HO CH OH KDO 出2-10LPS中三种独特的辅 在arma中,脂多糖分子中的O特异侧链可用灵敏的血清学方法加以鉴定。这在传染 病的诊断中有其重要意义,例如由此可对某传染病的传染源进行地理定位等。 外壁层中的蛋白质在革兰氏阴性细菌细胞壁的外壁层中,嵌有多种蛋白质,主要有:① 基质蛋白protein):例如在&.ad中的孔蛋白((porin)就是 一种研究得较多的基质蛋 白,孔蛋白是一种三聚体结构,每一个亚单位的分子量为36000。由三聚体结构构成的充水 孔道横跨外壁层,可通过分子量小于800一900的亲水性营养物质,例如糖类(尤其是双糖】 氨基酸、二肽、三肽、青霉素和无机离子等。它使外壁层具有分子筛的功能:②外壁蛋白 (outer membrane protein):是一类特异性的运送蛋白或受体,可把较大的分子,例如维生素B: 麦芽寡糖、核酸降解物以及铁离子薮合剂一铁色素(ferrichrome)或肠整合素 )等输送入细胞内:③脂蛋白(():种类很多,主要的是分子量为7200 的脂蛋白,其作用是使细胞壁的外壁层牢固地连接在由肽聚糖所组成的内壁层上, 周质空间(eriplasmic space)又称壁膜空间.指位于细胞壁与细胞漠之间的狭卒间隙 革兰氏阳性细菌与阴性细菌均有。内中含有多种蛋白质,例如蛋白酶、核酸等各种解聚酶, 运送某些物质进入细胞内的结合蛋白,以及趋化性的受体蛋白等。 革兰氏染色的机制通过以上对细菌细胞壁的详细分析,就为解释苹兰氏染色的机制提 供了较充分的基础。目前一般认为,革兰氏染色是基于细菌细胞壁特殊化学组分基础上的 种物理原因。通过初染和煤染操作后,在细菌细胞的膜或原生质体上染上了不溶于水的结晶 紫与碘的大分子复合物。革兰氏阳性细菌由于细胞壁较厚、肽聚糖含量较高和其分子交联度 较紧密,放在用乙醇洗脱时,肽聚糖网孔会因脱水而明显收缩,再加上它基本上不含类脂,故 乙醇处理不能在壁上溶出缝隙,因此,结晶紫与祺复合物仍牢牢阻留在其细胞壁内,使其呈 现紫色。反之,革兰氏阴性细菌因其壁薄、肽聚糖含量低和交联松散,故乙醇后,肽聚糖 网孔不易收缩,加上它的类脂含量高,所以当乙醇把类脂溶解后,在细胞壁上就会出现较大 的缝隙,这样,结晶紫与碘的复合物就极易被溶出细胞壁,因此,通过乙醇脱色后,细胞又 呈无色。这时,再经沙黄等红色染料进行复染,就使革兰氏阴性细菌获得了一层新的颜色 红色,而革兰氏阳性菌则仍呈紫色(实为紫中带红)。 (2)细胞膜与间体 细胞膜(cell me mbrane membrane),是紧贴在细胞壁内侧的一层由磷脂和蛋白质组成的柔软、富有弹性的半透性薄 36

膜。通过质壁分离、选择性染色、原生质体酸裂或电子显微镜观察等方法,可以证明细胞膜 的存在。 在电子显微镜下戏察时,细跑膜呈明显的双层结构一一在上下两暗色层间夹着一浅色的 中间层。这是因为,细胞膜的基本成分是由两层磷脂分子整齐地捧列而成的 每一磷脂分子由一个带正电荷且能溶于水的极性头(磷酸端)和一个不带电荷、不溶于 水的非极性尾(经端)所物成(图2-11)。极性头糊向膜的内外两个表面,虽亲水性:而非极 性的水尾(长链脂肪酸 ,其链长和饱和度与细菌的生长鼠度有关)则理藏在膜的内层,从 而形成一个醉脂双分子层。据目前所知,磷脂双分子层通常呈液态,不同的内嵌蛋白(穿过 全膜的、不对称地分布在膜的一烟或埋慧在隙脂双分子层内的)和外周蛋白可在磷脂双分子 层液体中作侧向运动,犹如漂浮在海洋中的冰山那样,这就是Singer和Nic (1972)提出 的细胞膜液态镶嵌模式的基本内容。有关漠蛋白在细胞膜中的分布和种类见图212。 -0-c9 V R-0- -0-c0w 0-H OH 甘袖3一酸 (方婆的路的) 图2山确脂的分子结构(函中的R基有多种形式,简如,R=一H(确脂酸),民■一:一GH: NH时(纳脂肤乙陈成):R=一CH2一GH一CO0(磷粉酰丝氨酸):R=一CH,一CH2一N+(H)《辆酯 分O服腕Q女服1服 wggO0O型 图2-12膜蛋白在生物离中的分和种癸 4内蛋白《不对称地分布于膜的一边点内嵌蛋白(穿过全):内蛋白(埋在脑双分子层 内》,《外简蛋白:,多牌复合体《由内数蛋白亚基和外圈蛋白亚基组成,穿过全膜,天.脂双分 细胞膜的功能为:①控制细胞内、 外的物质(营养物质和代谢废物)的运送 维持细胞内正常渗透压的屏难作用:③合成细跑壁各种组分(LS、肽聚糖、磷壁酸)和荚膜 等大分子的场所:④进行氧化酸化或光合弹酸化的产能基地:⑤许多酶(B半乳糖苷酶、有 关细胞壁和英膜的合成酶、ATP薛)和电子传递链组分的所在部位,⑧鞭毛的者生点和提供 其运动所需的能量等。 间体(mesosome)由细跑膜内熠形成的一一种管状、层状或赛状结构,一般位于细胞分裂 部位或其邻近,其功能主要是促进细胞间隔的形成并与遗传物质的复制及其相互分离有关 (图2-13)。在革兰氏阳性细菌如Bacils s6(枯草杆菌)、B.c时rm(地衣芽孢杆菌)、 Stre pucoc5 faecalis(粪链球黄)和ficrocuceus bdes(藤黄微球菊)中,间体较为明显
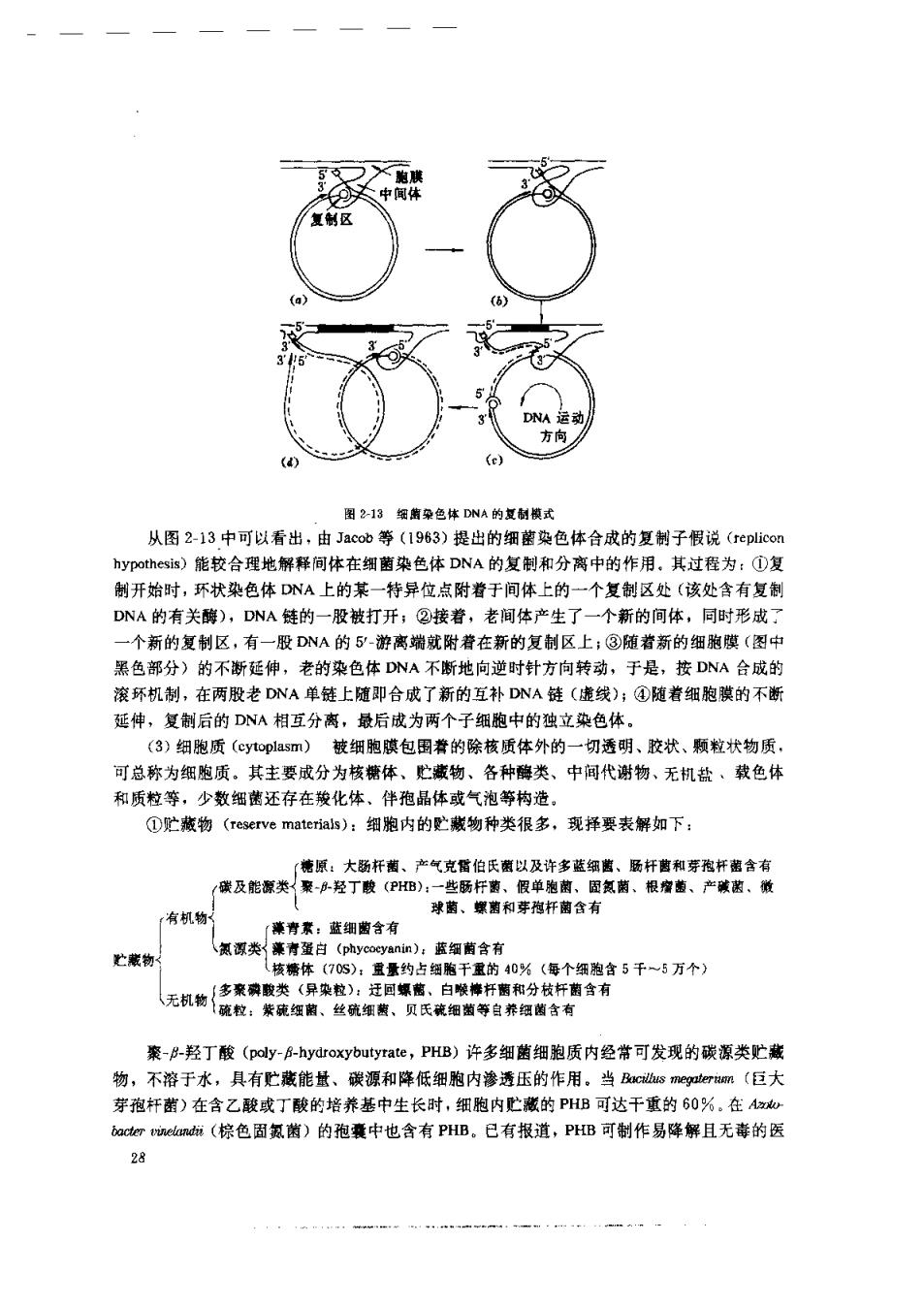
复制区 (a) (d) e 图213经的色体州A的制式 从图2-13中可以看出,由Jaco0等(1963)提出的细菌染色体合成的复树子假说(repi ypothesis)能较合理地解释间体在细菌染色体DNA的复制和分离中的作用,其过程为:①复 开始时,环状染色体DNA上的某一特异位点附着于间体上的一个复制区处(该处含有复制 DNA的有关酶),DNA链的一股被打开,②接着,老间体产生了 一个新的间体 同时形成 一个新的复制区,有一股DNA的5-游离端就附着在新的复制区上;③随着新的细胞膜(图中 黑色部分)的不断延伸,老的染色体DNA不断地向逆时针方向转动,于是,按DNA合成的 滚环机制,在两股老DNA单链上随即合成了新的互补DNA链(虚线);④随着细胞装的不断 延伸,复制后的DNA相互分离,最后成为两个子细胞中的独立染色体。 (3)细胞质(©ytoplasm)被细胞膜包围的除核质体外的一切透明、胶状、颗粒状物质, 可总称为细胞质。其主要成分为核糖体、贮藏物、各种酶类、中间代谢物、无机盐、载色体 和质粒等,少数细菌还存在羧化体、伴孢晶体或气泡等构造。 ①贮藏物(reserve materials):细胞内的贮戴物种类很多,现择要表解如下 德原:大肠杆葡、产气克猫伯氏菌以及许多蓝细菌、肠杆曹和芽孢杆菌含有 /碳及能原类装丹羟丁酸(PHB):一些所行菌、假单指前、图氮菌、根常整、声膜菌、散 有机物 球菌、螺菌和芽孢杆菌含有 贮紫物 A: 细曲 多豪碗酸类(异染老 无机物{统粒:紫成细潮、丝疏细费,贝氏使细菌等包养短篮含有 聚-B-羟丁酸(paly-hydroxybutyrate,PHB)许多细蓝细胞质内经常可发现的碳源类贮裁 物,不溶于水,具有贮藏能盘、碳源和降低细胞内渗透压的作用。当hs 芽孢杆菌)在含乙酸或丁酸的培养基中生长时,细胞内贮藏的PHB可达干重的60%,在 bacter rina城(棕色固氯菌)的孢囊中也含有PHB。已有报道,PHB可制作易降解且无毒的医 28