
第二章微生物的生理60学时) [内容提要]细菌菌体形成,涉及物质摄取、生物合成、聚合作用及组装四个步骤。菌体以均 等二分裂方式无性繁殖。细菌群体的生长曲线可分迟缓期、对数期、稳定期及衰亡期四个阶 段。大多数细菌可在人工培养基上生长。根据细菌对氧气和温度的需求差异,可将其分为需 氧菌、厌氧菌、兼性厌氧菌以及嗜温菌、嗜冷菌、嗜热菌等。由于细菌代谢的不同,以生化 试验用于细菌鉴定。细菌生存有赖于自身的调节机制,特别是细菌应激调节因子及其趋化性。 细菌与其他生物一样,有独立的生命活动,涉及复杂的新陈代谢。细菌的生理涉及细菌 的组分、营养要求、能量代谢、生物生长合成繁殖及基因调控等。本章将以细菌群体的生长 繁殖为主线,介绍细菌个体形成的代谢过程、群体的生长繁殖、细菌自身的调节机制以及工 人培养条件下细菌对营养的需求。 第一节细菌细胞的代谢过程 细菌的新陈代谢包括菌细胞的生物合成、能量供给、运动以及多达2000种化学反应表现 的各种活性。形成菌细胞的代谢过程可按其功能分为物质摄取、生物合成、聚合作用及组装 四个步骤(图2-1)
22 第二章 微生物的生理(6.0 学时) [内容提要] 细菌菌体形成,涉及物质摄取、生物合成、聚合作用及组装四个步骤。菌体以均 等二分裂方式无性繁殖。细菌群体的生长曲线可分迟缓期、对数期、稳定期及衰亡期四个阶 段。大多数细菌可在人工培养基上生长。根据细菌对氧气和温度的需求差异,可将其分为需 氧菌、厌氧菌、兼性厌氧菌以及嗜温菌、嗜冷菌、嗜热菌等。由于细菌代谢的不同,以生化 试验用于细菌鉴定。细菌生存有赖于自身的调节机制,特别是细菌应激调节因子及其趋化性。 细菌与其他生物一样,有独立的生命活动,涉及复杂的新陈代谢。细菌的生理涉及细菌 的组分、营养要求、能量代谢、生物生长合成繁殖及基因调控等。本章将以细菌群体的生长 繁殖为主线,介绍细菌个体形成的代谢过程、群体的生长繁殖、细菌自身的调节机制以及工 人培养条件下细菌对营养的需求。 第一节 细菌细胞的代谢过程 细菌的新陈代谢包括菌细胞的生物合成、能量供给、运动以及多达 2000 种化学反应表现 的各种活性。形成菌细胞的代谢过程可按其功能分为物质摄取、生物合成、聚合作用及组装 四个步骤(图 2-1)。 进入
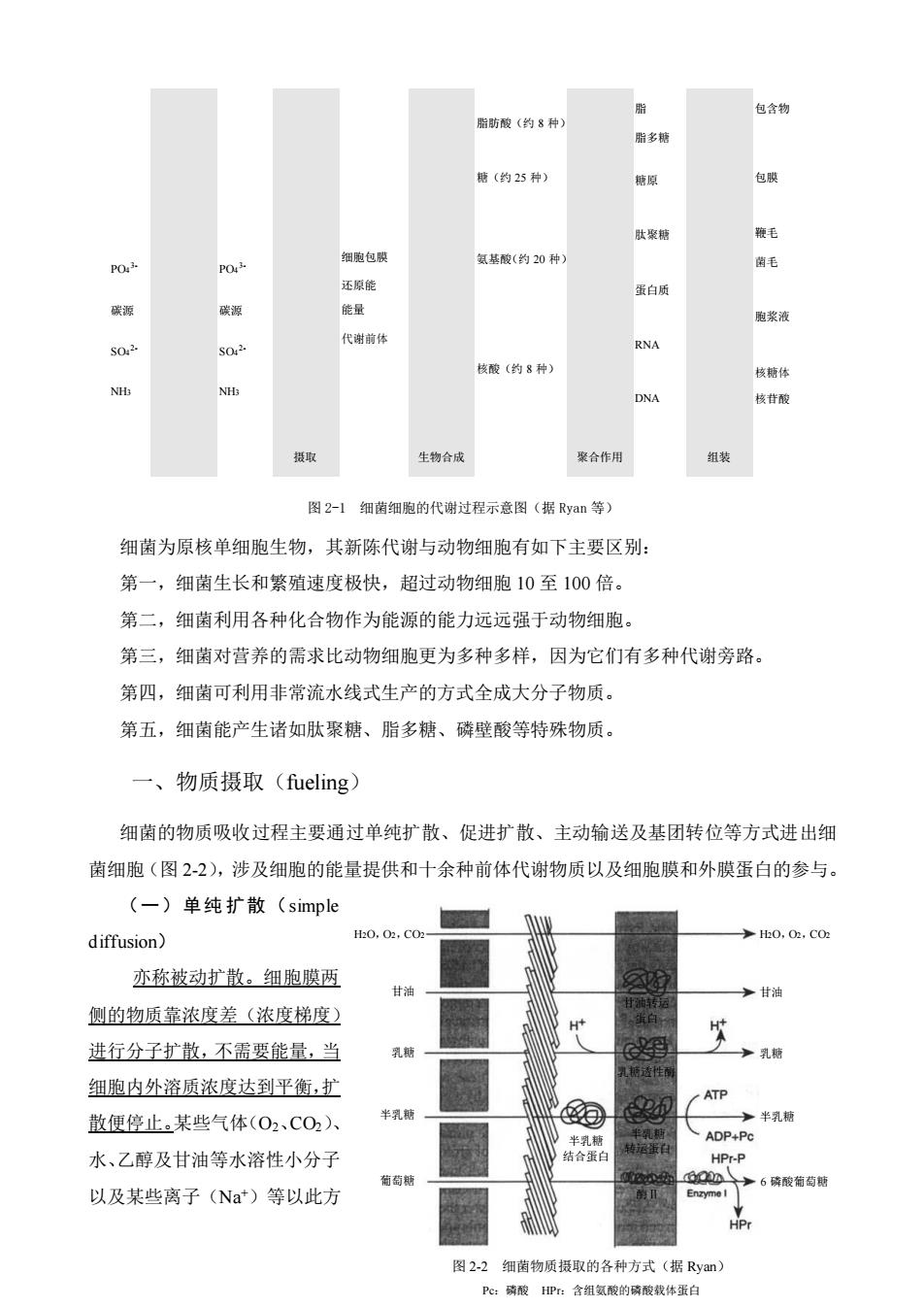
脂 脂防酸(约8种》 包含物 胎多 糖(约25种】 糖原 包 细飞包联 氢基酸(约20种 204 毛 还原能 蛋白质 碳源 绝量 胞浆 代瑞前 RNA 枝酸(约8种 H NH 世酸 生物合成 聚合作用 组装 图2-1细菌细胞的代谢过程示意图(据Ryan等) 细菌为原核单细胞生物,其新陈代谢与动物细胞有如下主要区别: 第一,细菌生长和繁殖速度极快,超过动物细胞10至100倍。 第二,细菌利用各种化合物作为能源的能力远远强于动物细胞。 第三,细菌对营养的需求比动物细胞更为多种多样,因为它们有多种代谢旁路。 第四,细菌可利用非常流水线式生产的方式全成大分子物质 第五,细菌能产生诸如肽聚糖、脂多糖、磷壁酸等特殊物质。 一、物质摄取(fueling 细菌的物质吸收过程主要通过单纯扩散、促进扩散、主动输送及基团转位等方式进出细 菌细胞(图2-2),涉及细胞的能量提供和十余种前体代谢物质以及细胞膜和外膜蛋白的参与。 (一)单纯扩散(simple diffusion) H0,0,C0 亦称被动扩散。细胞膜两 侧的物质靠浓度差(浓度梯度) 进行分子扩散,不需要能量,当 细胞内外溶质浓度达到平衡,扩 散便停止。某些气体(02,C02)入 水、乙醇及甘油等水溶性小分子 结合 面药 磷酸背 以及某些离子(Na*)等以此方 图22细南物质摄取的各种方式(据Rym) Pe:酸HP含组氢酸的酸找体蛋自
23 PO4 3- 碳源 SO4 2- NH3 PO4 3- 碳源 SO4 2- NH3 摄取 细胞包膜 还原能 能量 代谢前体 生物合成 脂肪酸(约 8 种) 糖(约 25 种) 氨基酸(约 20 种) 核酸(约 8 种) 聚合作用 脂 脂多糖 糖原 肽聚糖 蛋白质 RNA DNA 组装 包含物 包膜 鞭毛 菌毛 胞浆液 核糖体 核苷酸 图 2-1 细菌细胞的代谢过程示意图(据 Ryan 等) 细菌为原核单细胞生物,其新陈代谢与动物细胞有如下主要区别: 第一,细菌生长和繁殖速度极快,超过动物细胞 10 至 100 倍。 第二,细菌利用各种化合物作为能源的能力远远强于动物细胞。 第三,细菌对营养的需求比动物细胞更为多种多样,因为它们有多种代谢旁路。 第四,细菌可利用非常流水线式生产的方式全成大分子物质。 第五,细菌能产生诸如肽聚糖、脂多糖、磷壁酸等特殊物质。 一、物质摄取(fueling) 细菌的物质吸收过程主要通过单纯扩散、促进扩散、主动输送及基团转位等方式进出细 菌细胞(图 2-2),涉及细胞的能量提供和十余种前体代谢物质以及细胞膜和外膜蛋白的参与。 (一)单纯 扩散(simple diffusion) 亦称被动扩散。细胞膜两 侧的物质靠浓度差(浓度梯度) 进行分子扩散,不需要能量,当 细胞内外溶质浓度达到平衡,扩 散便停止。某些气体(O2、CO2)、 水、乙醇及甘油等水溶性小分子 以及某些离子(Na+)等以此方 图 2-2 细菌物质摄取的各种方式(据 Ryan) Pc:磷酸 HPr:含组氨酸的磷酸载体蛋白 H2O,O2,CO2 甘油 乳糖 半乳糖 葡萄糖 半乳糖 结合蛋白 H2O,O2,CO2 甘油 乳糖 半乳糖 6 磷酸葡萄糖 甘油转运 蛋白 乳糖透性酶 半乳糖 转运蛋白 酶Ⅱ

式运输。 (二)促进扩散(facilitated diffusion) 基些物质如糖或氨基酸与某些位于细胞膜的特异性载体蛋白(carrier protein)相结合, 而后将其转运至细胞内,这一过程具有特异性和选择性,也不需要能量。甘油进入大肠杆菌 等肠道菌的细胞内,厌氧菌吸收营养物质及排出代谢产物,均是通过这一方式。载体蛋白在 革兰氏阴性菌属于OMP,可能是通过构象改变而完成促进扩散的。 (三)主动输送(active transport) 是细菌吸收营养物质的一种主要方式,与促进扩散一样,需要特异性的载体蛋白,能将 特异性溶质逆浓度梯度“泵”入细胞,因此需要能量。 革兰氏阴性菌有两种主动输送的方式:休克敏感式(shock-.sensitive)及休克不敏感式 (shock-.insensitive)。在作渗透休克处理时,前者载体蛋白释放出所结合的可溶性分子如半乳 糖,通过细菌外膜的微孔进入细胞,该过程伴随ATP的水解。休克不敏感伴随H的跨膜,其 能量由获能的细胞膜的电子运输而获得,有特异性载体蛋白(透性酶)参与,乳糖的输送即 用此方式。 (四)基团转位(group translocation) 物质在运输的同时受到化学修饰,从而能源源不断输入细胞,此种方式在缺氢环境中最 为常见。其过程涉及从可溶物向分子的化学变化,需要特异性载体蛋白参与,需要能量。有 磷酸转移酶系统(phosphotransferase system)参与的葡萄糖磷酸化是个例证。 (五)金属离子的吸收 影响细菌生长的金属离子在动物体内极少以游离状态存在,以铁为例,它与铁结合蛋白 结合,如血清中的转铁蛋白及乳中的乳铁蛋白。细菌通过分泌载铁体攫取这些蛋白结合的 e3+,形成含铁鳌合物,而后通过特异的主动输送,F©3+进入菌体细胞。 载铁体(siderophore)是细菌在低铁条件下所产生的一类有 机化合物,分子量一般不超过1000,与F*有极强的亲合力。其合 成受铁调控,输送铁的功能与细菌外膜的受体有关。目前将载铁体 分为两种类型,一种为异羟肟盐类(hydroxamate),具有单个或两 个异羟肟酸功能团,气菌素(aerobactin)是其代表(图2-3),能抵 抗血清的灭活作用。另一种为酚盐类(phenolate),由2,3二羟基苯 CH-CH2-0B- 甲酸(2,3-DHS)与氨基酸偶联而成,肠菌素(enterobactin)是其代 图2-3大肠杆菌的气菌素 表,能被血清灭活。大肠杆菌具有这两种载铁体
24 式运输。 (二)促进扩散(facilitated diffusion) 某些物质如糖或氨基酸与某些位于细胞膜的特异性载体蛋白(carrier protein)相结合, 而后将其转运至细胞内,这一过程具有特异性和选择性,也不需要能量。甘油进入大肠杆菌 等肠道菌的细胞内,厌氧菌吸收营养物质及排出代谢产物,均是通过这一方式。载体蛋白在 革兰氏阴性菌属于 OMP,可能是通过构象改变而完成促进扩散的。 (三)主动输送(active transport) 是细菌吸收营养物质的一种主要方式,与促进扩散一样,需要特异性的载体蛋白,能将 特异性溶质逆浓度梯度“泵”入细胞,因此需要能量。 革兰氏阴性菌有两种主动输送的方式:休克敏感式(shock-sensitive)及休克不敏感式 (shock-insensitive)。在作渗透休克处理时,前者载体蛋白释放出所结合的可溶性分子如半乳 糖,通过细菌外膜的微孔进入细胞,该过程伴随 ATP 的水解。休克不敏感伴随 H+的跨膜,其 能量由获能的细胞膜的电子运输而获得,有特异性载体蛋白(透性酶)参与,乳糖的输送即 用此方式。 (四)基团转位(group translocation) 物质在运输的同时受到化学修饰,从而能源源不断输入细胞,此种方式在缺氧环境中最 为常见。其过程涉及从可溶物向分子的化学变化,需要特异性载体蛋白参与,需要能量。有 磷酸转移酶系统(phosphotransferase system)参与的葡萄糖磷酸化是个例证。 (五)金属离子的吸收 影响细菌生长的金属离子在动物体内极少以游离状态存在,以铁为例,它与铁结合蛋白 结合,如血清中的转铁蛋白及乳中的乳铁蛋白。细菌通过分泌载铁体攫取这些蛋白结合的 Fe3+,形成含铁螯合物,而后通过特异的主动输送,Fe3+进入菌体细胞。 载铁体(siderophore) 是细菌在低铁条件下所产生的一类有 机化合物,分子量一般不超过 1000,与 Fe3+有极强的亲合力。其合 成受铁调控,输送铁的功能与细菌外膜的受体有关。目前将载铁体 分为两种类型,一种为异羟肟盐类(hydroxamate),具有单个或两 个异羟肟酸功能团,气菌素(aerobactin)是其代表(图 2-3),能抵 抗血清的灭活作用。另一种为酚盐类(phenolate),由 2,3-二羟基苯 甲酸(2,3-DHS)与氨基酸偶联而成,肠菌素(enterobactin)是其代 表,能被血清灭活。大肠杆菌具有这两种载铁体。 O Fe3+ | O C=O | NH | Cyclo(CO—CH—CH2—O)3 — 图 2-3 大肠杆菌的气菌素

二、生物合成(biosynthesis) 吸收的各种前体代谢物通过代谢途径的网络,合成多种氨基酸、核苷酸、糖、脂肪酸及 其他合成大分子所需物质(图21)。此过程还需要碳的前体、还原的烟酰胺腺嘌吟二核苷酸 磷酸盐(NADPH)、ATP、氨基氨以及硫。不同种类细菌对营养的需要有所不同,又有其不 同的合成途径,据此可作为细菌实验室鉴定的重要指标, 三、聚合作用(polymerization) 细菌DNA的聚合作用称为复 制。其DNA复制从基因组的特定 起始部位开始,而后沿环状的染色 体DNA的复制叉(replication fork) 图24大肠杆菌染色体的复制义(据Madigan等) 部位,从5端到3'端双向进行(图2-4)。以半保留(semiconservative)方式复制,其DNA 双链解离,其中一条链作为母板合成互补链。另一条链则沿相反方向复制,最后由酶连接成 完成DNA链(图2-5)。细菌DNA复制的频率因细菌细胞的生长率而异,DNA链的延伸率 (elongation rate)则相对稳定。 某些抗生素以不同途径干扰细菌DNA的复制,例 复制起始部包 如新生霉素等抑制细菌DNA复制过程中所需的促旋 酶的活性。 (一)转录 1.细菌的转录特点:在某些方面与真核生物不同。 ①细菌可由同一个RNA聚合酶催化合成细菌的 mRNA、RNA及rRNA。该酶还可像真核生物那样利 用活化的ATP、GITP、CTP及UTP,并可作为母板合 成DNA互补链。②细菌mRNA不需要通过核膜转运 到胞浆,因此不需要聚A帽状结构,也不要特异的转 运方式,而且在mRNA合成早期直接与核糖体蛋白结 合形成多聚体。 2.RNA聚合酶:该酶是一个复杂的大分子,由a2 BB1G亚单位组成。o亚单位可与特异的DNA序列 即启动子(promoter)结合。细菌RNA聚合酶一般具 国25大超开他D到度过际金
25 二、生物合成(biosynthesis) 吸收的各种前体代谢物通过代谢途径的网络,合成多种氨基酸、核苷酸、糖、脂肪酸及 其他合成大分子所需物质(图 2-1)。此过程还需要碳的前体、还原的烟酰胺腺嘌呤二核苷酸 磷酸盐(NADPH)、ATP、氨基氮以及硫。不同种类细菌对营养的需要有所不同,又有其不 同的合成途径,据此可作为细菌实验室鉴定的重要指标。 三、聚合作用(polymerization) 细菌 DNA 的聚合作用称为复 制。其 DNA 复制从基因组的特定 起始部位开始,而后沿环状的染色 体 DNA 的复制叉(replication fork) 部位,从 5’端到 3’端双向进行(图 2-4)。以半保留(semiconservative)方式复制,其 DNA 双链解离,其中一条链作为母板合成互补链。另一条链则沿相反方向复制,最后由酶连接成 完成 DNA 链(图 2-5)。细菌 DNA 复制的频率因细菌细胞的生长率而异,DNA 链的延伸率 (elongation rate)则相对稳定。 某些抗生素以不同途径干扰细菌 DNA 的复制,例 如新生霉素等抑制细菌 DNA 复制过程中所需的促旋 酶的活性。 (一)转录 1. 细菌的转录特点:在某些方面与真核生物不同。 ①细菌可由同一个 RNA 聚合酶催化合成细菌的 mRNA、tRNA 及 rRNA。该酶还可像真核生物那样利 用活化的 ATP、GTP、CTP 及 UTP,并可作为母板合 成 DNA 互补链。②细菌 mRNA 不需要通过核膜转运 到胞浆,因此不需要聚 A 帽状结构,也不要特异的转 运方式,而且在 mRNA 合成早期直接与核糖体蛋白结 合形成多聚体。 2. RNA 聚合酶:该酶是一个复杂的大分子,由α2 ββ1σ亚单位组成。σ亚单位可与特异的 DNA 序列 即启动子(promoter)结合。细菌 RNA 聚合酶一般具 图 2-4 大肠杆菌染色体的复制叉(据 Madigan 等) 复制叉 新合成的 DNA 复制起始部位 图2-5 大肠杆菌染色体 DNA 的复制过程示意图 (据 Madigan 等)

有一个以上的σ亚单位,可识别不同的启动子,从而激活相关的基因。RNA聚合酶是利福平 类药物的靶分子,其可阻断转录的起始。 3.RNA的加工修饰:像真核细胞一样,RNA大分子的前体被核酸酶等加工、修饰,而后 产生稳定的RNA分子如RNA、rRNA等。 (二)翻译 也称转译,是蛋白质合成过程。20种氨基酸激活后与相应的tRNA结合,形成的氨基酰 RNA通过某些可溶性蛋白因子与核糖体结合。在此过程中,氨基酸根据mRNA上的密码子 序列聚合成多肽。tRNA释放出氨基酸后,自身也从核糖体上脱落,并进入下一轮氨基酰循环。 多种抗生素以细菌翻译过程有抑制作用,氯霉素、林可莓素和红霉素等抑制核糖体大亚 基,四环素、链霉素和壮观霉素等抑制小亚基,还有一些作用于核链孢酸(fusidicacid)等可 溶性蛋白因子,后者参与多肽的合成。 以上过程在原核细胞和真核细胞基本一致,但存在一些差别。细菌核糖体较小,结构也较简单。细菌 的mRNA不需要经过加工和转运,为大的顺反子,包含多个基因,可指导多个多肽链的合成。细菌RNA 聚合酶合成mRNA的速率为55个核苷酸/秒(37℃),核糖体合成多肽链的速率为18个氨基酸/秒。因此 细菌中mRNA的翻译与DNA的转录不仅同时进行,而且两者速率也相等(每秒55个核苷酸/侮个密码子 3个核苷酸=每秒18个氨基酸),这意味着核糖体在mRNA链上移动速率与RNA聚合酶合成mRNA的速 率相等。 细菌的上述翻译特性决定了细菌高效合成过程。细菌胞质内充满多聚核糖体。每个核糖体几乎都最大 限度地发挥作用。细菌生长越快,需要合成蛋白质用的核糖体就越多。在丰富培养基上生长的大肠杆菌 一半以上菌体由核糖体和其它翻译组分构成。 四、组装(assembly) 细菌细胞结构的组装有两种方式:自我组装(self-assembly,又名自我凝聚),及指导组 装(guided assembly,又名特异机制)。自我组装可在体外试管内完成,鞭毛及核糖体即采用 此种方式。细菌表面膜结构则只能依赖指导组装来完成,过程尚不完全清楚,涉及蛋白质的 分泌,跨膜运输及载体分子如细菌萜醇等。 杆菌肽或万古霉素可干扰细菌细胞成份组装所需的载体的功能,多粘菌素可影响细胞膜 的组装。 第二节细菌的生长繁殖 细菌生长繁殖分别体现在个体和群体两方面。 26
26 有一个以上的σ亚单位,可识别不同的启动子,从而激活相关的基因。RNA 聚合酶是利福平 类药物的靶分子,其可阻断转录的起始。 3. RNA 的加工修饰:像真核细胞一样,RNA 大分子的前体被核酸酶等加工、修饰,而后 产生稳定的 RNA 分子如 tRNA、rRNA 等。 (二)翻译 也称转译,是蛋白质合成过程。20 种氨基酸激活后与相应的 tRNA 结合,形成的氨基酰 -tRNA 通过某些可溶性蛋白因子与核糖体结合。在此过程中,氨基酸根据 mRNA 上的密码子 序列聚合成多肽。tRNA 释放出氨基酸后,自身也从核糖体上脱落,并进入下一轮氨基酰循环。 多种抗生素以细菌翻译过程有抑制作用,氯霉素、林可霉素和红霉素等抑制核糖体大亚 基,四环素、链霉素和壮观霉素等抑制小亚基,还有一些作用于核链孢酸(fusidic acid)等可 溶性蛋白因子,后者参与多肽的合成。 以上过程在原核细胞和真核细胞基本一致,但存在一些差别。细菌核糖体较小,结构也较简单。细菌 的 mRNA 不需要经过加工和转运,为大的顺反子,包含多个基因,可指导多个多肽链的合成。细菌 RNA 聚合酶合成 mRNA 的速率为 55 个核苷酸/秒(37℃),核糖体合成多肽链的速率为 18 个氨基酸/秒。因此 细菌中 mRNA 的翻译与 DNA 的转录不仅同时进行,而且两者速率也相等(每秒 55 个核苷酸/每个密码子 3 个核苷酸=每秒 18 个氨基酸),这意味着核糖体在 mRNA 链上移动速率与 RNA 聚合酶合成 mRNA 的速 率相等。 细菌的上述翻译特性决定了细菌高效合成过程。细菌胞质内充满多聚核糖体。每个核糖体几乎都最大 限度地发挥作用。细菌生长越快,需要合成蛋白质用的核糖体就越多。在丰富培养基上生长的大肠杆菌, 一半以上菌体由核糖体和其它翻译组分构成。 四、组装(assembly) 细菌细胞结构的组装有两种方式:自我组装(self-assembly,又名自我凝聚),及指导组 装(guided assembly,又名特异机制)。自我组装可在体外试管内完成,鞭毛及核糖体即采用 此种方式。细菌表面膜结构则只能依赖指导组装来完成,过程尚不完全清楚,涉及蛋白质的 分泌,跨膜运输及载体分子如细菌萜醇等。 杆菌肽或万古霉素可干扰细菌细胞成份组装所需的载体的功能,多粘菌素可影响细胞膜 的组装。 第二节 细菌的生长繁殖 细菌生长繁殖分别体现在个体和群体两方面