
四、生态分布和意义 (一)生态分布 多毛类动物种类多,分布于所有海洋生境中,对纬度、深度和底质都表现出明显的特 异性。以沙蚕科动物为例,从潮间带上部至深海带都有分布。如褐片阔沙蚕Platynereis dumerili从潮间带至48s0m(北大西洋)片环带沙蚕Nereis2onaa从水深8m(青岛胶州湾)至 3899m(大西洋)都有分布。大多数种类为沿岸浅海种,最大深度182m(东海陆架区)。从生态 类型可知,有的沙蚕生活在潮间带,各种类型的潮间带岩岸、石沼、珊瑚礁、红树林,软质 的各种底质(软泥、泥砂、砂质泥和细砂)都有沙蚕栖息,潮下带至深海的各种底质也几乎 都可以采取沙蚕。有的种类则生活于河口及淡水水域。从海水,咸淡水(半盐水)至淡水(远 离河口的广州或南京)都有分布。我国有4种沙蚕能在淡水中生活,其中有1种长须单叶沙 蚕Namalycastis仅柄于淡水,为纯淡水种:有13种能生活在咸淡水的河口地带,均系广盐 种:另有9种能适应较高盐度的变化栖于海水,如日本刺沙蚕能适应Q.93一30的盐度变化。 (二)经济意义 1.有益方面在海洋生态系的物质循环和能量流动中,多毛类是食物链中的一个重要环 节,无论成虫或幼虫均可作为经济甲壳类和经济鱼类(不仅底层鱼,也包括一些中、上层鱼) 的饵料,具有重要经济意义: 多毛类幼虫属阶段性浮游生物,在海洋浮游生物中占一定数量,有些种如双管阁沙蚕 和日本刺沙蚕的担轮幼虫、后担轮幼虫和疣足幼虫都营浮游生活,是对虾幼体和稚鱼的优良 饵料。目前我国沿海许多养虾池在早春日本刺沙蚕繁殖时期开闸纳潮,沙蚕幼虫随之而入, 数量极大,成为对虾养殖期间极重要的饵料动物。 多毛类如沙蚕科的成虫营底栖生活,在岩岸石块下、石缝中、海藻丛中及珊瑚礁周丛 生物中均为优势种。很多种类在软相的底质上分布非常稠密,因此在底栖动物的生物量中占 优势。据统计,在34种鱼的消化道中,有多毛类者就占14种之多,象底栖鱼中的鲽鱼和 鳄类,沙蚕等多毛类动物在其胃含物中的总数量可达50%一80%。 沙蚕、齿吻沙蚕Nephtyidae、吻沙蚕、矶沙蚕Eunicidae等科的许多种类,在生死季 节多有二次变态现象,此时,其雌、雄沙蚕体在月光刺激下从底内云集、浮游于海面,这一 习性会引起鱼类的集群。我国沿海的经济鱼类如鲐、参鱼群,都是在仔鱼、小虾和浮游动物 集中的海域形成鱼场的。当沙蚕群浮时,由于数量集中,体内充满了生殖腺,其营养物质高 度地集中,使浮游生物的质和量急剧递增,为鱼类极易捕食和喜好。鱼类成群地追逐、捕食, 形成良好的捕食区。据报道,当红鮭润游到堪察加时,正值绿沙蚕的异沙蚕体群浮云集海面 之际,由此可见群浮沙蚕与酒游索饵鱼群之间的密切关系。总之,多毛类对渔场分布、渔业 资源的状况、鱼类对产卵场的选择都有较密切的关系。 多毛类可做为海洋生态环境的指示种。如小头虫科Capitellidae生活于沿岸,吞咽型食 性,近厌氧呼吸,对有机物有很强的耐受力,每年3一5月份可大量采到。又如海稚虫科的
四、生态分布和意义 (一)生态分布 多毛类动物种类多,分布于所有海洋生境中,对纬度、深度和底质都表现出明显的特 异性。以沙蚕科动物为例,从潮间带上部至深海带都有分布。如褐片阔沙蚕 Platynereis dumerilii 从潮间带至 4850m(北大西洋);环带沙蚕 Nereis zonata 从水深 8m(青岛胶州湾)至 3899m(大西洋)都有分布。大多数种类为沿岸浅海种,最大深度 182m(东海陆架区)。从生态 类型可知,有的沙蚕生活在潮间带,各种类型的潮间带岩岸、石沼、珊瑚礁、红树林,软质 的各种底质(软泥、泥砂、砂质泥和细砂)都有沙蚕栖息,潮下带至深海的各种底质也几乎 都可以采取沙蚕。有的种类则生活于河口及淡水水域。从海水,咸淡水(半盐水)至淡水(远 离河口的广州或南京)都有分布。我国有4种沙蚕能在淡水中生活,其中有1种长须单叶沙 蚕 Namalycastis 仅栖于淡水,为纯淡水种;有 13 种能生活在咸淡水的河口地带,均系广盐 种;另有 9 种能适应较高盐度的变化栖于海水,如日本刺沙蚕能适应 0.93─30 的盐度变化。 (二)经济意义 1.有益方面 在海洋生态系的物质循环和能量流动中,多毛类是食物链中的一个重要环 节,无论成虫或幼虫均可作为经济甲壳类和经济鱼类(不仅底层鱼,也包括一些中、上层鱼) 的饵料,具有重要经济意义: 多毛类幼虫属阶段性浮游生物,在海洋浮游生物中占一定数量,有些种如双管阔沙蚕 和日本刺沙蚕的担轮幼虫、后担轮幼虫和疣足幼虫都营浮游生活,是对虾幼体和稚鱼的优良 饵料。目前我国沿海许多养虾池在早春日本刺沙蚕繁殖时期开闸纳潮,沙蚕幼虫随之而入, 数量极大,成为对虾养殖期间极重要的饵料动物。 多毛类如沙蚕科的成虫营底栖生活,在岩岸石块下、石缝中、海藻丛中及珊瑚礁周丛 生物中均为优势种。很多种类在软相的底质上分布非常稠密,因此在底栖动物的生物量中占 优势。据统计,在34种鱼的消化道中,有多毛类者就占14种之多,象底栖鱼中的鲽鱼和 鳕类,沙蚕等多毛类动物在其胃含物中的总数量可达 50%─80%。 沙蚕、齿吻沙蚕 Nephtyidae、吻沙蚕、矶沙蚕 Eunicidae 等科的许多种类,在生死季 节多有二次变态现象,此时,其雌、雄沙蚕体在月光刺激下从底内云集、浮游于海面,这一 习性会引起鱼类的集群。我国沿海的经济鱼类如鲐、参鱼群,都是在仔鱼、小虾和浮游动物 集中的海域形成鱼场的。当沙蚕群浮时,由于数量集中,体内充满了生殖腺,其营养物质高 度地集中,使浮游生物的质和量急剧递增,为鱼类极易捕食和喜好。鱼类成群地追逐、捕食, 形成良好的捕食区。据报道,当红鲑洄游到堪察加时,正值绿沙蚕的异沙蚕体群浮云集海面 之际,由此可见群浮沙蚕与洄游索饵鱼群之间的密切关系。总之,多毛类对渔场分布、渔业 资源的状况、鱼类对产卵场的选择都有较密切的关系。 多毛类可做为海洋生态环境的指示种。如小头虫科 Capitellidae 生活于沿岸,吞咽型食 性,近厌氧呼吸,对有机物有很强的耐受力,每年 3─5 月份可大量采到。又如海稚虫科的

奇异稚齿虫Paraparionospio pinnata是11一12月份离岸稍远的有机物指标生物。根据它们 在生物群落中出现的优占度,可以说明海洋底质的污染程度。再有,沙蚕科很多广盐性种如 日本刺沙蚕、琥珀沙蚕、疣吻沙蚕常栖于河口附近,是天然的污水监测者。据调查,日本刺 沙蚕在严重污染条件下形态发生畸形:疣足退化、刚毛落脱。可见沙蚕不论幼虫还是成虫, 其发育和形态都能反映水域遭受污染的程度。 温度是生物地理分布的一个重要限制因素,从地理分布可知,有的多毛类是仅生活在热 带和亚热带的暖水种,如双齿围沙蚕,岩虫Marphysa sanguinea等:有的则是只生活于冷水 环境的冷水种,如齿吻沙蚕Nephtyidae类等。它们的存在,可做为环境冷暖程度的指标。 例如,我国黄海北部是冷水种分布的南界,在黄海,大部分冷水种多栖于潮下带,大部分暖 水种则栖于潮间带,这一事实说明黄海潮下带比潮间带有着更为稳定的冷水条件,也说明黄 海北部和中部是北太平洋温带和西太平洋热带区之间的过渡带。 此外,管柄多毛类的古生态学研究,在指示地质成因,寻找石油资源方面提供了旁证, 这也是目前新发展的课题之一 海洋污损生物(附者生物)对水产养殖业和渔业的危害是多方面的。在附着的多毛环 虫中以石灰质栖管的龙介虫(石灰虫)和螺旋虫危害最大。龙介虫多附着于岩石、贝类、珊 瑚、海藻叶片和其它硬物上。据研究,内刺盘管虫是我国北方的优势种:而华美盘管虫我国 南北方极常见,它们危害贝类养殖,可使牡蛎大量减产,在我国北方,7月份螺旋虫栖总密 度可达每平方米两万余个,其它如缨鳃虫、蛰龙介以及鳞沙蚕Aphroditidae、沙蚕、裂虫等 都是附着生物群落中的成员。 2有害方面另一类多毛类环虫亦属该除之列:在珍珠贝养殖中,被称为黑壳病或黑心 脏病的,其病源就是凿贝而居的才女虫属Po口,才女虫能分泌腐蚀贝壳的物质,在蚀透 贝壳钻孔而居的过程中导致珍珠贝被细菌感染而发生脓肿渍疡,致使珍珠贝的死亡率高达 60%一80%,危害甚大。最近的研究还表明,首次在我国纪录的利氏才女虫Pgm每年春 季大量出现于辽河三角洲的蟹苗池中,对河蟹Eriocheir sinensis蚤状幼体和褶皱臂尾轮虫 Brachionus plicatilis的危害十分严重。 还有一些广盐性的疣吻沙蚕和多齿围沙蚕Perinereis nuntia能在低盐和淡水中生活,在 南方常栖于沿海稻田,啮食稻根:危害农作物,实为农业上的害虫。 沙蜀Arenicolidae是温带河口地区泥滩的一种优势生物,据报道,在一英里的海滩上, 它们每年能把19001泥沙搬到表面,大约两年就能把50一60cm厚的泥沙滩耕翻一遍。类似 例子很多,有人统计,在一个一英里的沙岸,有一亿五千万条海蛹Opheliidae,湿重约70t, 它能使沙中14000:有机物周年循环。可见多毛环虫对海弹软底质中的物质循环具有深刻影 响
奇异稚齿虫 Paraparionospio pinnata 是 11─12 月份离岸稍远的有机物指标生物。根据它们 在生物群落中出现的优占度,可以说明海洋底质的污染程度。再有,沙蚕科很多广盐性种如 日本刺沙蚕、琥珀沙蚕、疣吻沙蚕常栖于河口附近,是天然的污水监测者。据调查,日本刺 沙蚕在严重污染条件下形态发生畸形:疣足退化、刚毛落脱。可见沙蚕不论幼虫还是成虫, 其发育和形态都能反映水域遭受污染的程度。 温度是生物地理分布的一个重要限制因素,从地理分布可知,有的多毛类是仅生活在热 带和亚热带的暖水种,如双齿围沙蚕,岩虫 Marphysa sanguinea 等;有的则是只生活于冷水 环境的冷水种,如齿吻沙蚕 Nephtyidae 类等。它们的存在,可做为环境冷暖程度的指标。 例如,我国黄海北部是冷水种分布的南界,在黄海,大部分冷水种多栖于潮下带,大部分暖 水种则栖于潮间带,这一事实说明黄海潮下带比潮间带有着更为稳定的冷水条件,也说明黄 海北部和中部是北太平洋温带和西太平洋热带区之间的过渡带。 此外,管栖多毛类的古生态学研究,在指示地质成因,寻找石油资源方面提供了旁证, 这也是目前新发展的课题之一。 海洋污损生物(附着生物)对水产养殖业和渔业的危害是多方面的。在附着的多毛环 虫中以石灰质栖管的龙介虫(石灰虫)和螺旋虫危害最大。龙介虫多附着于岩石、贝类、珊 瑚、海藻叶片和其它硬物上。据研究,内刺盘管虫是我国北方的优势种;而华美盘管虫我国 南北方极常见,它们危害贝类养殖,可使牡蛎大量减产,在我国北方,7月份螺旋虫栖息密 度可达每平方米两万余个,其它如缨鳃虫、蛰龙介以及鳞沙蚕 Aphroditidae、沙蚕、裂虫等 都是附着生物群落中的成员。 2.有害方面 另一类多毛类环虫亦属该除之列:在珍珠贝养殖中,被称为黑壳病或黑心 脏病的,其病源就是凿贝而居的才女虫属 Polydora,才女虫能分泌腐蚀贝壳的物质,在蚀透 贝壳钻孔而居的过程中导致珍珠贝被细菌感染而发生脓肿溃疡,致使珍珠贝的死亡率高达 60%─80%,危害甚大。最近的研究还表明,首次在我国纪录的利氏才女虫 P. ligni 每年春 季大量出现于辽河三角洲的蟹苗池中,对河蟹 Eriocheir sinensis 蚤状幼体和褶皱臂尾轮虫 Brachionus plicatilis 的危害十分严重。 还有一些广盐性的疣吻沙蚕和多齿围沙蚕 Perinereis nuntia 能在低盐和淡水中生活,在 南方常栖于沿海稻田,啮食稻根;危害农作物,实为农业上的害虫。 沙蜀 Arenicolidae 是温带河口地区泥滩的一种优势生物,据报道,在一英里的海滩上, 它们每年能把 1900t 泥沙搬到表面,大约两年就能把 50─60cm厚的泥沙滩耕翻一遍。类似 例子很多,有人统计,在一个一英里的沙岸,有一亿五千万条海蛹 Opheliidae,湿重约 70t, 它能使沙中 14000t 有机物周年循环。可见多毛环虫对海洋软底质中的物质循环具有深刻影 响
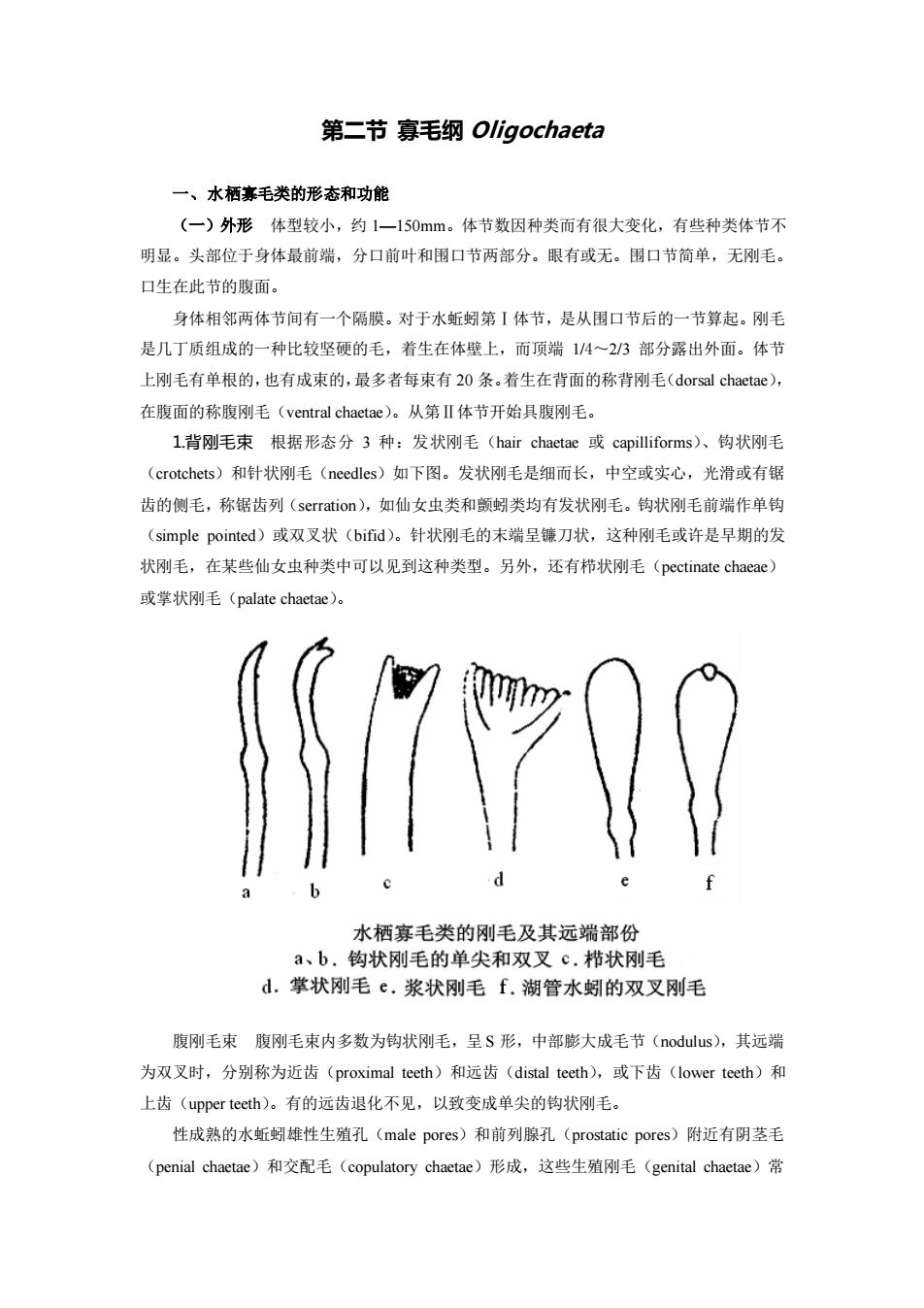
第二节寡毛纲Oligochaeta 一、水栖毛类的形态和功能 (一)外形体型较小,约1一150mm。体节数因种类而有很大变化,有些种类体节不 明显。头部位于身体最前端,分口前叶和围口节两部分。眼有或无。围口节简单,无刚毛。 口生在此节的腹面。 身体相邻两体节间有一个隔膜。对于水蚯蚓第I体节,是从围口节后的一节算起。刚毛 是几丁质组成的一种比较坚硬的毛,若生在体壁上,而顶端14~23部分露出外面。体节 上刚毛有单根的,也有成束的,最多者每束有20条。着生在背面的称背刚毛(dorsal chaetae), 在腹面的称腹刚毛(ventral chaetae)。从第Ⅱ体节开始具腹刚毛。 L背刚毛束根据形态分3种:发状刚毛(hair chaetae或capilliforms)、钩状刚毛 (crotchets)和针状刚毛(needles)如下图。发状刚毛是细而长,中空或实心,光滑或有锯 齿的侧毛,称锯齿列(serration),如仙女虫类和颤蚓类均有发状刚毛。钩状刚毛前端作单钩 (simple pointed)或双叉状(bifid)。针状刚毛的末端呈镰刀状,这种刚毛或许是早期的发 状刚毛,在某些仙女虫种类中可以见到这种类型。另外,还有栉状刚毛(pectinate chacae》 或掌状刚毛(palate chaetae), 770 水栖寡毛类的刚毛及其远端部份 a、b,钩状刚毛的单尖和双叉c.栉状刚毛 d.掌状刚毛e.浆状刚毛f.湖管水蚓的双叉刚毛 腹刚毛束腹刚毛束内多数为钩状刚毛,呈S形,中部膨大成毛节(nodulus),其远端 为双叉时,分别称为近齿(proximal teeth)和远齿(distal teeth),或下齿(lower teeth)和 上齿(upper tecth).。有的远齿退化不见,以致变成单尖的钩状刚毛。 性成熟的水蚯蚓雄性生殖孔(male pores)和前列腺孔(prostatic pores)附近有阴茎毛 (penial chaetae)和交配毛(copulatory chaetae)形成,这些生殖刚毛(genital chaetae)常
第二节 寡毛纲 Oligochaeta 一、水栖寡毛类的形态和功能 (一)外形 体型较小,约 1—150mm。体节数因种类而有很大变化,有些种类体节不 明显。头部位于身体最前端,分口前叶和围口节两部分。眼有或无。围口节简单,无刚毛。 口生在此节的腹面。 身体相邻两体节间有一个隔膜。对于水蚯蚓第Ⅰ体节,是从围口节后的一节算起。刚毛 是几丁质组成的一种比较坚硬的毛,着生在体壁上,而顶端 1/4~2/3 部分露出外面。体节 上刚毛有单根的,也有成束的,最多者每束有 20 条。着生在背面的称背刚毛(dorsal chaetae), 在腹面的称腹刚毛(ventral chaetae)。从第Ⅱ体节开始具腹刚毛。 1.背刚毛束 根据形态分 3 种:发状刚毛(hair chaetae 或 capilliforms)、钩状刚毛 (crotchets)和针状刚毛(needles)如下图。发状刚毛是细而长,中空或实心,光滑或有锯 齿的侧毛,称锯齿列(serration),如仙女虫类和颤蚓类均有发状刚毛。钩状刚毛前端作单钩 (simple pointed)或双叉状(bifid)。针状刚毛的末端呈镰刀状,这种刚毛或许是早期的发 状刚毛,在某些仙女虫种类中可以见到这种类型。另外,还有栉状刚毛(pectinate chaeae) 或掌状刚毛(palate chaetae)。 腹刚毛束 腹刚毛束内多数为钩状刚毛,呈 S 形,中部膨大成毛节(nodulus),其远端 为双叉时,分别称为近齿(proximal teeth)和远齿(distal teeth),或下齿(lower teeth)和 上齿(upper teeth)。有的远齿退化不见,以致变成单尖的钩状刚毛。 性成熟的水蚯蚓雄性生殖孔(male pores)和前列腺孔(prostatic pores)附近有阴茎毛 (penial chaetae)和交配毛(copulatory chaetae)形成,这些生殖刚毛(genital chaetae)常

常两根在一起。阴茎毛和交配毛在形状和大小上变化很大(如下图),很少有相同、重复的 形状,因此其形态可以作为分类的依据。 几种水蚯蚓阴茎毛的远端 a.矛坞蚜oodwardiella hastate b.马什兰双胃蚓 Eudichogaster matheranensis c,正蚓科交配毛横切 示沟与脊d.短毛仙女虫Naidium breviseta的阴茎 毛,具两个长尖叉 (二)主要器官的结构与功能 1.呼吸器宫无专门的呼吸器官,通过身体表面及皮下的微血管进行气体交换,有些 种类用鰓呼吸或肠壁进行呼吸。水蚯蚓的血液为红色或黄色,不具红血球,以血红脘溶于血 浆内。呼吸作用一般利用皮肤下的微血管交换气体。如颤蚓T万br和水丝蝴是通过身体表 面积的增大和加快颤动次数,促使周围水流更新来调节呼吸的。它们的尾部突出淤泥,体前 端则埋入泥中。在正常情况下尾部有节奏地摆动,进行平静的呼吸。颜蚓类喜生活在氧饱和 率10一60%的水中,当水中含氧量不足时,则拉长身体并增加尾部的颜动速度。 有些种类在身体前端或尾端具有由皮肤形成的特殊的鰓,以此进行呼吸。在水栖寡毛类 中,鰓呼吸仅见于仙女虫科中尾盘虫属Dem、管盘虫属Aulophorus、头鰓虫属Branchiodrilus 和甑蚓科的苏氏尾鳃蚓ranchiura sowrerbyri等。尾盘虫和盘管虫的身体末端都有一个浅杯 状结构,称鳃窝(branchial foss)。鰓窝略朝后上方。肛门位于其中。从其内壁伸出的称真 鳃(true gills),,从边缘伸出的称副鳃(accessory gills)。头鳃蚓属的鳃是按节成队排列的 位于体前端,始自第各节的背侧面,向后渐细小,终止于体后端。苏氏尾鳃蚓的鳃沿着背 腹中线按节排列成两纵行,共50-140对,其长度几乎与体宽相等,前端的鰓小而不显。生 活时,鳃有规律地从后向前摆动,促进了蚓体后面的运动。 2.生殖系统雌雄同体。生殖腺集中在前段体节内,雄性器官包括精巢、精巢囊、输精 管。输精管外端通雄孔。雌性生殖器包括卵巢囊,输卵管,受精囊(对外有开口的受精囊孔)。 输卵管内端有卵漏斗,外端通雌孔。水蚯蚓异体受精交配时,两个体前端以腹面相靠合
常两根在一起。阴茎毛和交配毛在形状和大小上变化很大(如下图),很少有相同、重复的 形状,因此其形态可以作为分类的依据。 (二)主要器官的结构与功能 1.呼吸器官 无专门的呼吸器官,通过身体表面及皮下的微血管进行气体交换,有些 种类用鳃呼吸或肠壁进行呼吸。水蚯蚓的血液为红色或黄色,不具红血球,以血红脘溶于血 浆内。呼吸作用一般利用皮肤下的微血管交换气体。如颤蚓 Tubifex 和水丝蚓是通过身体表 面积的增大和加快颤动次数,促使周围水流更新来调节呼吸的。它们的尾部突出淤泥,体前 端则埋入泥中。在正常情况下尾部有节奏地摆动,进行平静的呼吸。颤蚓类喜生活在氧饱和 率 10—60%的水中,当水中含氧量不足时,则拉长身体并增加尾部的颤动速度。 有些种类在身体前端或尾端具有由皮肤形成的特殊的鳃,以此进行呼吸。在水栖寡毛类 中,鳃呼吸仅见于仙女虫科中尾盘虫属 Dero、管盘虫属 Aulophorus、头鳃虫属 Branchiodrilus 和颤蚓科的苏氏尾鳃蚓 Vranchiura sowerbyri 等。尾盘虫和盘管虫的身体末端都有一个浅杯 状结构,称鳃窝(branchial fossa)。鳃窝略朝后上方。肛门位于其中。从其内壁伸出的称真 鳃(true gills),从边缘伸出的称副鳃(accessory gills)。头鳃蚓属的鳃是按节成队排列的, 位于体前端,始自第Ⅵ各节的背侧面,向后渐细小,终止于体后端。苏氏尾鳃蚓的鳃沿着背、 腹中线按节排列成两纵行,共 50-140 对,其长度几乎与体宽相等,前端的鳃小而不显。生 活时,鳃有规律地从后向前摆动,促进了蚓体后面的运动。 2.生殖系统 雌雄同体。生殖腺集中在前段体节内,雄性器官包括精巢、精巢囊、输精 管。输精管外端通雄孔。雌性生殖器包括卵巢囊,输卵管,受精囊(对外有开口的受精囊孔)。 输卵管内端有卵漏斗,外端通雌孔。 水蚯蚓异体受精交配时,两个体前端以腹面相靠合
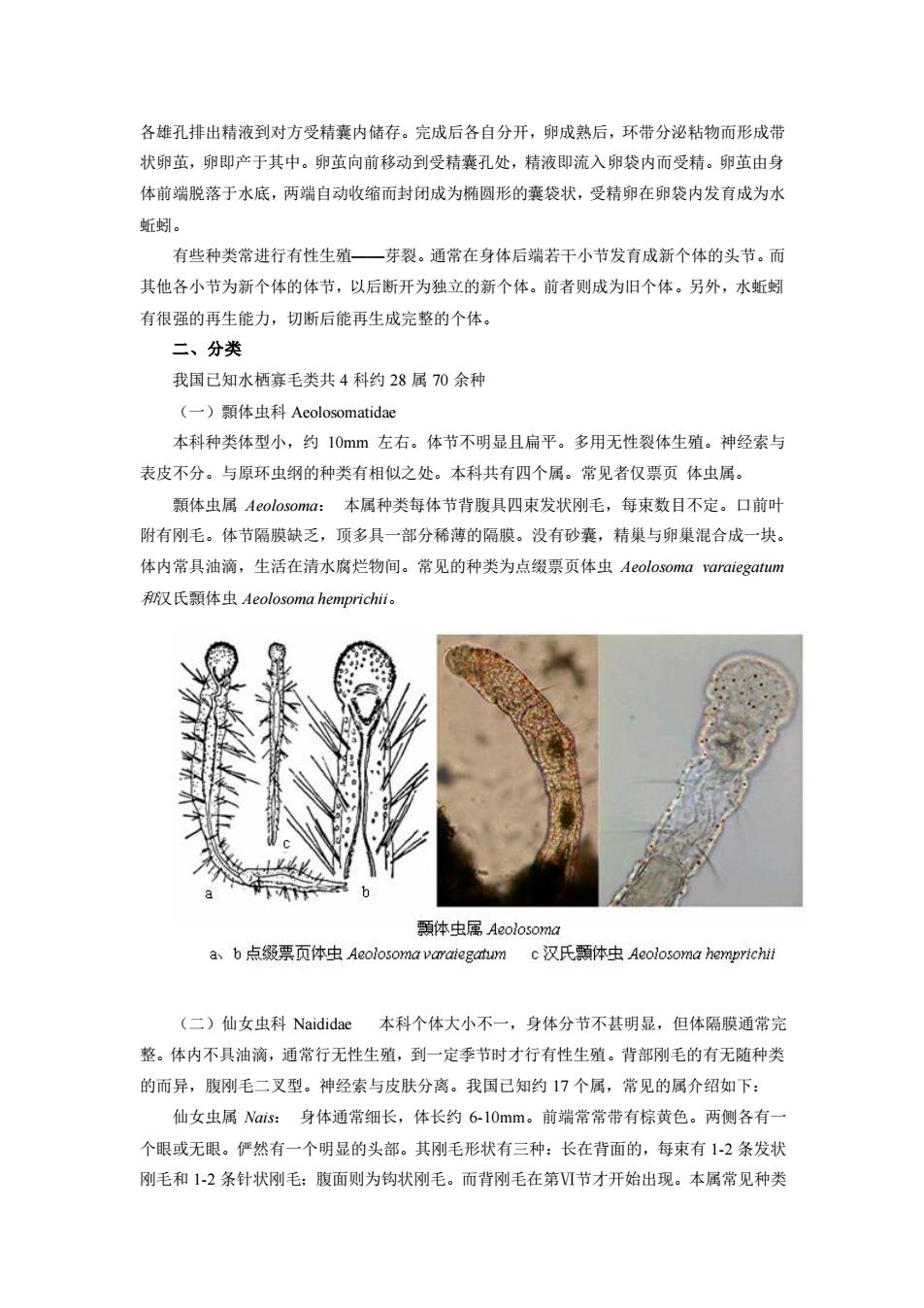
各雄孔排出精液到对方受精囊内储存。完成后各自分开,卵成熟后,环带分泌粘物而形成带 状卵茧,卵即产于其中。卵茧向前移动到受精囊孔处,精液即流入卵袋内而受精。卵茧由身 体前端脱落于水底,两端自动收缩而封闭成为椭圆形的囊袋状,受精卵在卵袋内发有成为水 斯蚓 有些种类常进行有性生殖一芽裂。通常在身体后端若干小节发育成新个体的头节。而 其他各小节为新个体的体节,以后断开为独立的新个体。前者则成为旧个体。另外,水蚯蚓 有很强的再生能力,切断后能再生成完整的个体。 二、分类 我国已知水栖寡毛类共4科约28属70余种 (一)领体虫科Aeolosomatidae 本科种类体型小,约10mm左右。体节不明显且扁平。多用无性裂体生殖。神经索与 表皮不分。与原环虫纲的种类有相似之处。本科共有四个属。常见者仅票页体虫属。 颗体虫属Aeolosoma: 本属种类每体节背腹具四束发状刚毛,每束数目不定。口前 附有刚毛。体节隔膜缺乏,顶多具一部分稀薄的隔膜。没有砂囊,精巢与卵巢混合成一块。 体内常具油滴,生活在清水腐烂物间。常见的种类为点缀票页体虫Aeolosoma1 araiegatum 氏短体虫Aeolosoma hemprichii。 顯体虫属Aeolosoma a、b点级票页体虫Aeolosoma varategatum c汉氏雨体中Aeolosoma hemprichit (二)仙女虫科Naididae本科个体大小不一,身体分节不甚明显,但体隔膜通常完 整。体内不具油滴,通常行无性生殖,到一定季节时才行有性生殖。背部刚毛的有无随种类 的而异,腹刚毛二叉型。神经索与皮肤分离。我国己知约17个属,常见的属介绍如下: 仙女虫属Nas:身体通常细长,体长约6-10mm。前端常常带有棕黄色。两侧各有 个眼或无眼。俨然有一个明显的头部。其刚毛形状有三种:长在背面的,每束有12条发状 刚毛和1-2条针状刚毛:腹面则为钩状刚毛。而背刚毛在第Ⅵ节才开始出现。本属常见种类
各雄孔排出精液到对方受精囊内储存。完成后各自分开,卵成熟后,环带分泌粘物而形成带 状卵茧,卵即产于其中。卵茧向前移动到受精囊孔处,精液即流入卵袋内而受精。卵茧由身 体前端脱落于水底,两端自动收缩而封闭成为椭圆形的囊袋状,受精卵在卵袋内发育成为水 蚯蚓。 有些种类常进行有性生殖——芽裂。通常在身体后端若干小节发育成新个体的头节。而 其他各小节为新个体的体节,以后断开为独立的新个体。前者则成为旧个体。另外,水蚯蚓 有很强的再生能力,切断后能再生成完整的个体。 二、分类 我国已知水栖寡毛类共 4 科约 28 属 70 余种 (一)顠体虫科 Aeolosomatidae 本科种类体型小,约 10mm 左右。体节不明显且扁平。多用无性裂体生殖。神经索与 表皮不分。与原环虫纲的种类有相似之处。本科共有四个属。常见者仅票页 体虫属。 顠体虫属 Aeolosoma: 本属种类每体节背腹具四束发状刚毛,每束数目不定。口前叶 附有刚毛。体节隔膜缺乏,顶多具一部分稀薄的隔膜。没有砂囊,精巢与卵巢混合成一块。 体内常具油滴,生活在清水腐烂物间。常见的种类为点缀票页体虫 Aeolosoma varaiegatum 和汉氏顠体虫 Aeolosoma hemprichii。 (二)仙女虫科 Naididae 本科个体大小不一,身体分节不甚明显,但体隔膜通常完 整。体内不具油滴,通常行无性生殖,到一定季节时才行有性生殖。背部刚毛的有无随种类 的而异,腹刚毛二叉型。神经索与皮肤分离。我国已知约 17 个属,常见的属介绍如下: 仙女虫属 Nais: 身体通常细长,体长约 6-10mm。前端常常带有棕黄色。两侧各有一 个眼或无眼。俨然有一个明显的头部。其刚毛形状有三种:长在背面的,每束有 1-2 条发状 刚毛和 1-2 条针状刚毛;腹面则为钩状刚毛。而背刚毛在第Ⅵ节才开始出现。本属常见种类