
利氏才女虫P.1igni A.体前部背面观B.体前端侧面观C.第5刚节足刺刚毛D.第5刚节的伴随刚毛 E有巾毛状背刚毛R另 种有巾背刚毛G.两齿腹巾钩刚毛H体后端背面观 龙介虫科Serpulidae具石灰质栖管,体对称,又名石灰虫。头部口前叶不明显,常 与围口节愈合形成漏斗状的鳃冠(触手冠)围绕着口,其上具很多鳃丝(触手丝),具呼吸 和激动水流滤食之功。鰓丝常具两排小羽枝(鰓羽枝),其背中线的一个鳃丝末端可变成能 封闭壳口的壳盖。躯干部可分为两区:胸区至少5个刚节,具胸膜,第1刚节多扩展成外翻 的领(有建管腺体)。胸区的背刚毛通常为具翅毛状刚毛或枪刺状或鳍刺状,腹刚毛为齿片 状(梳状)。腹区具很多体节,其刚毛的分节与胸区相反,背刚毛为齿片,腹刚毛为翅状或 刺叭状等。 龙介虫与缨鳃虫都具栖管。但龙介虫的栖管是石灰质的,无触角,常有壳盖,而缨鳃虫 栖管为角质颧泥沙膜质,有触角,无特化的壳盖。 石灰虫管多附者在岩石和其它硬物或珊瑚上,是一类极其重要的污损生物。如盘管虫属 等喜附者在浮标、船底、养殖设施或养殖贝类的壳表面。有壳盖的种类一般生活在潮间带, 无壳盖的种类生活在潮下带。如龙介虫属。 龙介虫属Serpula壳盖只有一层,漏斗状其边缘有锯齿,壳柄光滑。领刚毛毛状和枪 刺状,无弯刀刚毛。腹部刚毛为刺叭状,齿片刿毛有数个小齿。7个胸刚节。如龙介虫 S.vermicularis,柄管的切面为圆形。管表面有5一7个纵脊。为世界性种,我国沿海潮 间带、湖下带均有分布。是世界性主要污损生物之一
龙介虫科 Serpulidae 具石灰质栖管,体对称,又名石灰虫。头部口前叶不明显,常 与围口节愈合形成漏斗状的鳃冠(触手冠)围绕着口,其上具很多鳃丝(触手丝),具呼吸 和激动水流滤食之功。鳃丝常具两排小羽枝(鳃羽枝),其背中线的一个鳃丝末端可变成能 封闭壳口的壳盖。躯干部可分为两区:胸区至少5个刚节,具胸膜,第1刚节多扩展成外翻 的领(有建管腺体)。胸区的背刚毛通常为具翅毛状刚毛或枪刺状或鳍刺状,腹刚毛为齿片 状(梳状)。腹区具很多体节,其刚毛的分节与胸区相反,背刚毛为齿片,腹刚毛为翅状或 刺叭状等。 龙介虫与缨鳃虫都具栖管。但龙介虫的栖管是石灰质的,无触角,常有壳盖,而缨鳃虫 栖管为角质颧泥沙膜质,有触角,无特化的壳盖。 石灰虫管多附着在岩石和其它硬物或珊瑚上,是一类极其重要的污损生物。如盘管虫属 等喜附着在浮标、船底、养殖设施或养殖贝类的壳表面。有壳盖的种类一般生活在潮间带, 无壳盖的种类生活在潮下带。如龙介虫属。 龙介虫属 Serpula 壳盖只有一层,漏斗状其边缘有锯齿,壳柄光滑。领刚毛毛状和枪 刺状,无弯刀刚毛。腹部刚毛为刺叭状,齿片刿毛有数个小齿。7个胸刚节。如龙介虫 S.vermicularis ,栖管的切面为圆形。管表面有5─7个纵脊。为世界性种,我国沿海潮 间带、潮下带均有分布。是世界性主要污损生物之一
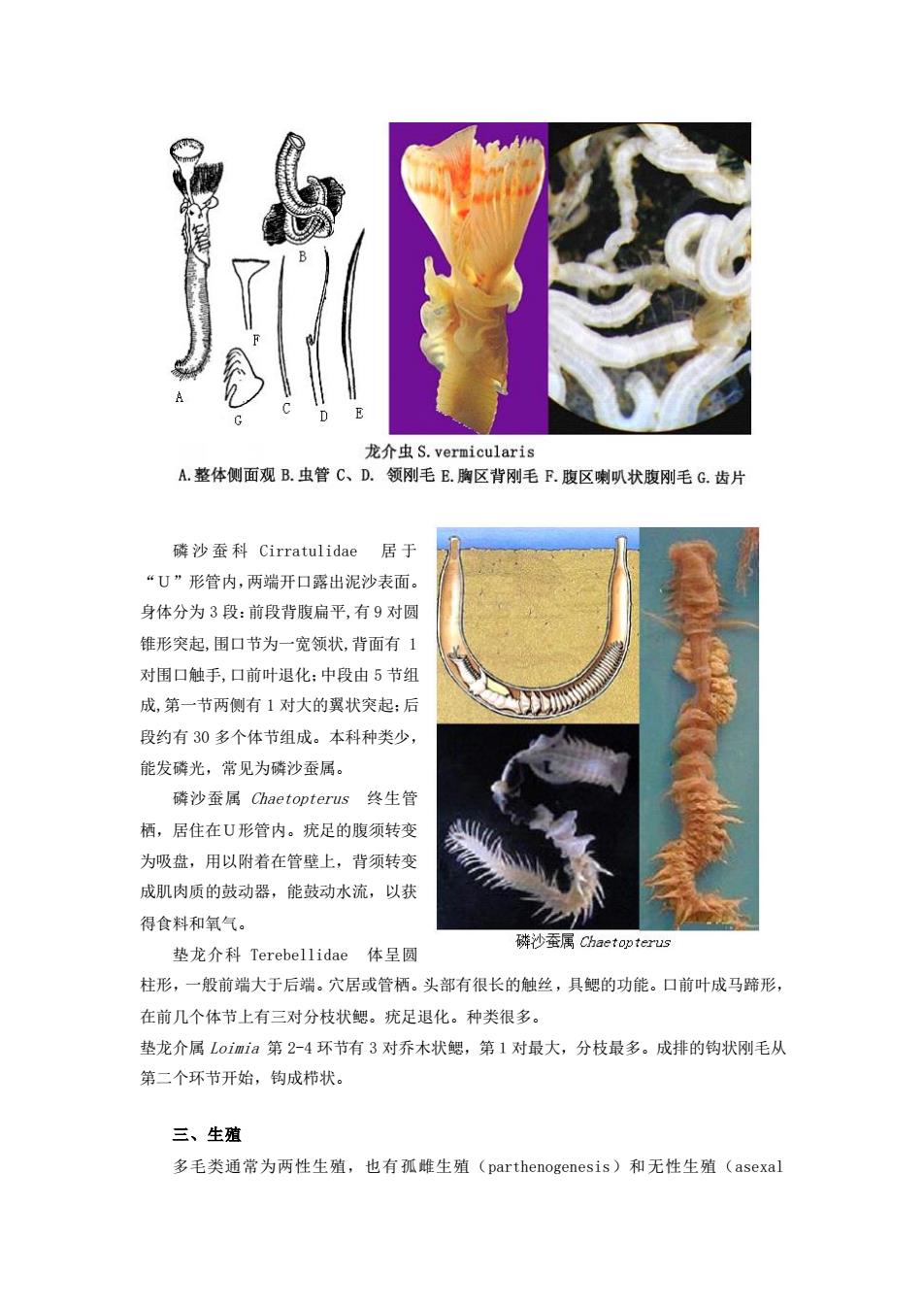
龙介虫S.vermicularis A.整体侧面观B.虫管C、D.领刚毛E胸区背刚毛R腹区喇叭状腹刚毛G.齿片 磷沙蚕科Cirratulidae居于 “U”形管内,两端开口露出泥沙表面。 身体分为3段:前段背腹扁平,有9对圆 锥形突起,围口节为一宽领状,背面有1 对围口触手,口前叶退化:中段由5节组 成,第一节两侧有1对大的翼状突起:后 段约有30多个体节组成。本科种类少, 能发磷光,常见为磷沙蚕属。 磷沙蚕属Chaetopterus终生管 栖,居住在U形管内。疣足的腹须转变 为吸盘,用以附着在管壁上,背须转变 成肌肉质的鼓动器,能鼓动水流,以获 得食料和氧气。 垫龙介科Terebellidae体呈圆 磷沙蚕属Chactopterus 柱形,一般前端大于后端。穴居或管栖。头部有很长的触丝,具鳃的功能。口前叶成马蹄形, 在前几个体节上有三对分枝状鳃。疣足退化。种类很多。 垫龙介属Loimia第2-4环节有3对乔木状鳃,第1对最大,分枝最多。成排的钩状刚毛从 第二个环节开始,钩成栉状。 三、生殖 多毛类通常为两性生殖,也有孤雌生殖(parthenogenesis)和无性生殖(asexal
磷沙蚕科 Cirratulidae 居 于 “U”形管内,两端开口露出泥沙表面。 身体分为 3 段:前段背腹扁平,有 9 对圆 锥形突起,围口节为一宽领状,背面有 1 对围口触手,口前叶退化;中段由 5 节组 成,第一节两侧有 1 对大的翼状突起;后 段约有 30 多个体节组成。本科种类少, 能发磷光,常见为磷沙蚕属。 磷沙蚕属 Chaetopterus 终生管 栖,居住在U形管内。疣足的腹须转变 为吸盘,用以附着在管壁上,背须转变 成肌肉质的鼓动器,能鼓动水流,以获 得食料和氧气。 垫龙介科 Terebellidae 体呈圆 柱形,一般前端大于后端。穴居或管栖。头部有很长的触丝,具鳃的功能。口前叶成马蹄形, 在前几个体节上有三对分枝状鳃。疣足退化。种类很多。 垫龙介属 Loimia 第 2-4 环节有 3 对乔木状鳃,第1对最大,分枝最多。成排的钩状刚毛从 第二个环节开始,钩成栉状。 三、生殖 多毛类通常为两性生殖,也有孤雌生殖(parthenogenesis)和无性生殖(asexal
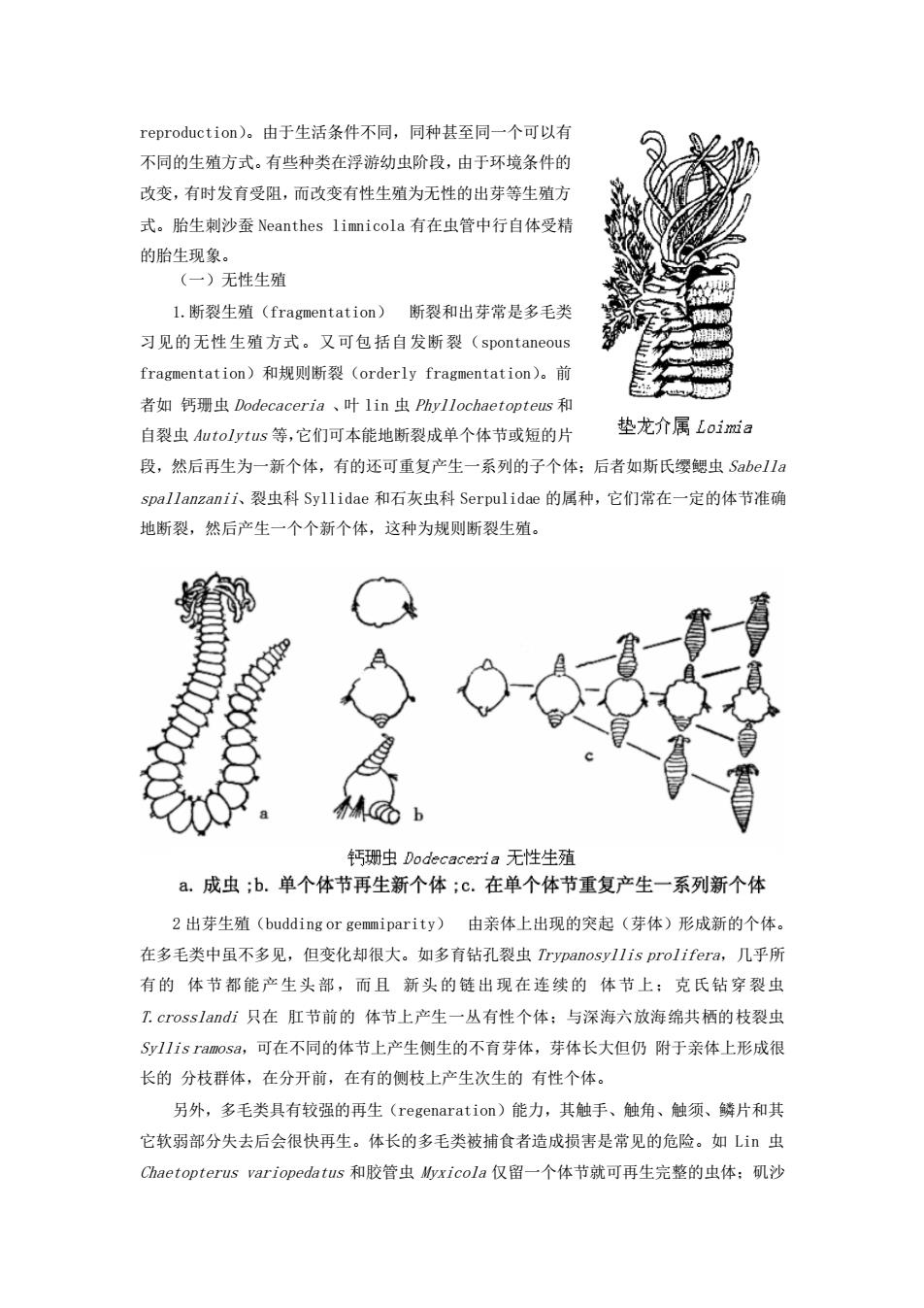
reproduction)。由于生活条件不同,同种甚至同一个可以有 不同的生殖方式。有些种类在浮游幼虫阶段,由于环境条件的 改变,有时发育受阻,而改变有性生殖为无性的出芽等生殖方 式。胎生刺沙蚕Neanthes limnicola有在虫管中行自体受精 的胎生现象。 (一)无性生殖 L.断裂生殖(fragmentation)断裂和出芽常是多毛类 习见的无性生殖方式。又可包括自发断裂(spontaneous fragmentation)和规则断裂(orderly fragmentation)。前 者如钙珊虫Dodecaceria、叶lin虫Phyllochaetopteus和 自裂虫A加tolytus等,它们可本能地断裂成单个体节或短的片 垫龙介属Loia 段,然后再生为一新个体,有的还可重复产生一系列的子个体:后者如斯氏缨鰓虫Sabella spallanzanii、裂虫科Syllidae和石灰虫科Serpulidae的属种,它们常在一定的体节准确 地断裂,然后产生一个个新个体,这种为规则断裂生殖 人M上 钙珊虫Dodecaceria无性生殖 a.成虫;b.单个体节再生新个体;c.在单个体节重复产生一系列新个体 2出芽生殖(budding or gemmiparity)由亲体上出现的突起(芽体)形成新的个体。 在多毛类中虽不多见,但变化却很大。如多育钻孔裂虫Trvpanosvllis pro/ifera,几乎所 有的体节都能产生头部,而且新头的链出现在连续的体节上:克氏钻穿裂虫 1.crosslandi只在肛节前的体节上产生一丛有性个体;与深海六放海绵共栖的枝裂虫 Sy11 is ramosa,可在不同的体节上产生侧生的不有芽体,芽体长大但仍附于亲体上形成很 长的分枝群体,在分开前,在有的侧枝上产生次生的有性个体。 另外,多毛类具有较强的再生(regenaration)能力,其触手、触角、触须、鳞片和其 它软弱部分失去后会很快再生。体长的多毛类被捕食者造成损害是常见的危险。如L虫 Chaetopterus variopedatus和胶管虫xicola仅留一个体节就可再生完整的虫体;矶沙
reproduction)。由于生活条件不同,同种甚至同一个可以有 不同的生殖方式。有些种类在浮游幼虫阶段,由于环境条件的 改变,有时发育受阻,而改变有性生殖为无性的出芽等生殖方 式。胎生刺沙蚕 Neanthes limnicola 有在虫管中行自体受精 的胎生现象。 (一)无性生殖 1.断裂生殖(fragmentation) 断裂和出芽常是多毛类 习见的无性生殖方式。又可包括自发断裂(spontaneous fragmentation)和规则断裂(orderly fragmentation)。前 者如 钙珊虫 Dodecaceria 、叶 lin 虫 Phyllochaetopteus 和 自裂虫 Autolytus 等,它们可本能地断裂成单个体节或短的片 段,然后再生为一新个体,有的还可重复产生一系列的子个体;后者如斯氏缨鳃虫 Sabella spallanzanii、裂虫科 Syllidae 和石灰虫科 Serpulidae 的属种,它们常在一定的体节准确 地断裂,然后产生一个个新个体,这种为规则断裂生殖。 2 出芽生殖(budding or gemmiparity) 由亲体上出现的突起(芽体)形成新的个体。 在多毛类中虽不多见,但变化却很大。如多育钻孔裂虫 Trypanosyllis prolifera,几乎所 有的 体节都能产生头部,而且 新头的链出现在连续的 体节上;克氏钻穿裂虫 T.crosslandi 只在 肛节前的 体节上产生一丛有性个体;与深海六放海绵共栖的枝裂虫 Syllis ramosa,可在不同的体节上产生侧生的不育芽体,芽体长大但仍 附于亲体上形成很 长的 分枝群体,在分开前,在有的侧枝上产生次生的 有性个体。 另外,多毛类具有较强的再生(regenaration)能力,其触手、触角、触须、鳞片和其 它软弱部分失去后会很快再生。体长的多毛类被捕食者造成损害是常见的危险。如 Lin 虫 Chaetopterus variopedatus 和胶管虫 Myxicola 仅留一个体节就可再生完整的虫体;矶沙
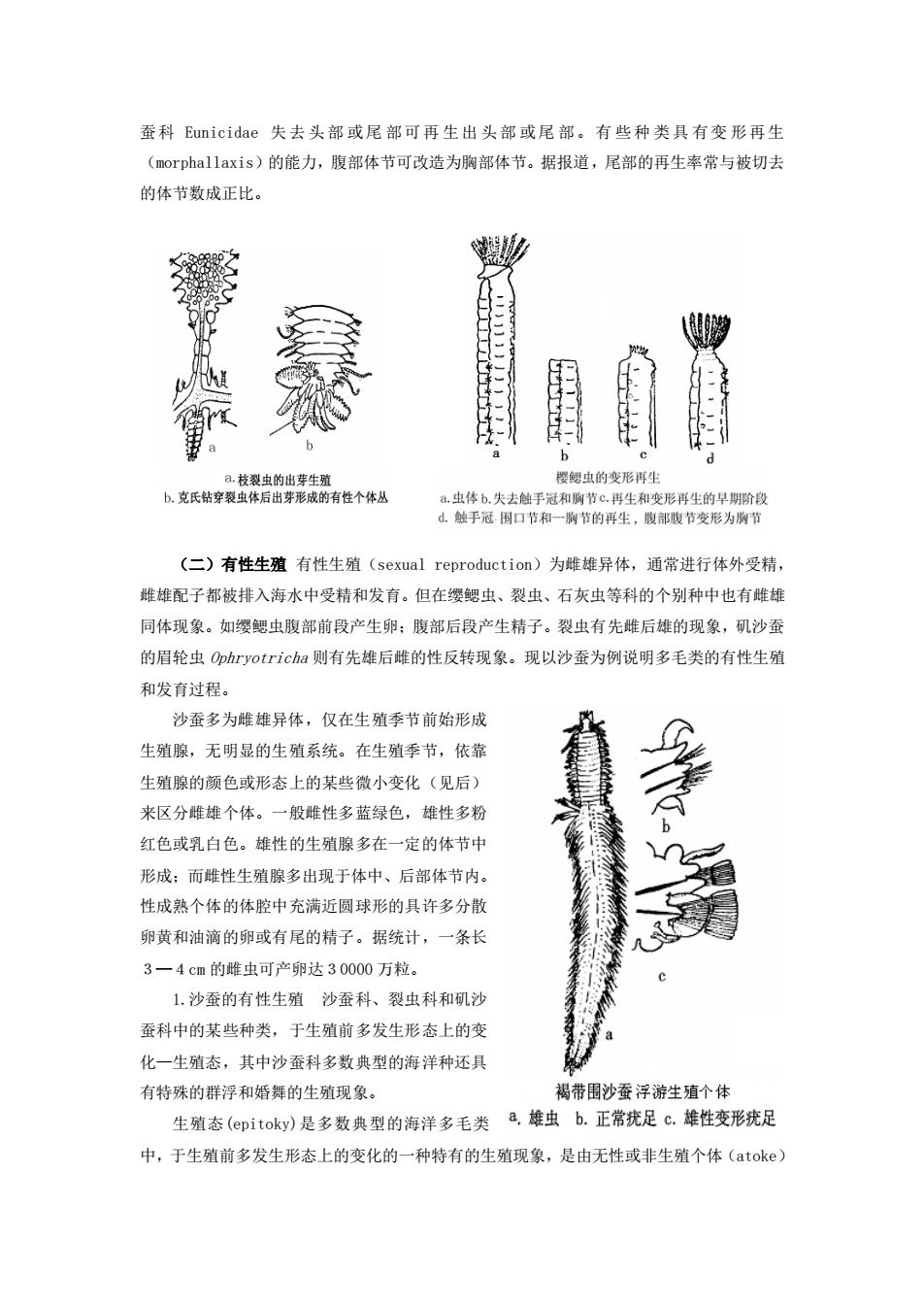
蚕科Cunicidae失去头部或尾部可再生出头部或尾部。有些种类具有变形再生 (morphallaxis)的能力,腹部体节可改造为胸部体节。据报道,尾部的再生率常与被切去 的体节数成正比, 1 a.枝虫的出芽生驼 樱鰓虫的变形再生 b.克氏钻穿表虫体后出芽形成的有性个体丛 a虫体b.失去触手冠和胸节c再生和变形再生的早期阶段 ,触手冠围口节和一刷节的再生,腹酸节变形为节 (二)有性生殖有性生殖(sexual reproduction)为雌雄异体,通常进行体外受精, 雌雄配子都被排入海水中受精和发有。但在缨鳃虫、裂虫、石灰虫等科的个别种中也有雌雄 同体现象。如缨鳃虫腹部前段产生卵:腹部后段产生精子。裂虫有先雌后雄的现象,矶沙蚕 的眉轮虫phryotricha则有先雄后雌的性反转现象。现以沙蚕为例说明多毛类的有性生殖 和发育过程。 沙蚕多为雌雄异体,仅在生殖季节前始形成 生殖腺,无明显的生殖系统。在生殖季节,依靠 生殖腺的颜色或形态上的某些微小变化(见后)》 来区分雌雄个体。一般雌性多蓝绿色,雄性多粉 红色或乳白色。雄性的生殖腺多在一定的体节中 形成:而雌性生殖腺多出现于体中、后部体节内。 性成熟个体的体腔中充满近圆球形的具许多分散 卵黄和油滴的卵或有尾的精子。据统计,一条长 3一4cm的雌虫可产卵达30000万粒。 1.沙蚕的有性生殖沙蚕科、裂虫科和乳沙 蚕科中的某些种类,于生殖前多发生形态上的变 化一生殖态,其中沙蚕科多数典型的海洋种还具 有特殊的群浮和婚舞的生殖现象。 褐带围沙蚕浮游生殖个体 生殖态(epitoky)是多数典型的海洋多毛类a,雄虫b.正常疣足c.雄性变形疣足 中,于生殖前多发生形态上的变化的一种特有的生殖现象,是由无性或非生殖个体(atoke)
蚕科 Eunicidae 失去 头部 或尾 部可再 生出 头部 或尾 部。有 些种 类具 有变 形再生 (morphallaxis)的能力,腹部体节可改造为胸部体节。据报道,尾部的再生率常与被切去 的体节数成正比。 (二)有性生殖 有性生殖(sexual reproduction)为雌雄异体,通常进行体外受精, 雌雄配子都被排入海水中受精和发育。但在缨鳃虫、裂虫、石灰虫等科的个别种中也有雌雄 同体现象。如缨鳃虫腹部前段产生卵;腹部后段产生精子。裂虫有先雌后雄的现象,矶沙蚕 的眉轮虫 Ophryotricha 则有先雄后雌的性反转现象。现以沙蚕为例说明多毛类的有性生殖 和发育过程。 沙蚕多为雌雄异体,仅在生殖季节前始形成 生殖腺,无明显的生殖系统。在生殖季节,依靠 生殖腺的颜色或形态上的某些微小变化(见后) 来区分雌雄个体。一般雌性多蓝绿色,雄性多粉 红色或乳白色。雄性的生殖腺多在一定的体节中 形成;而雌性生殖腺多出现于体中、后部体节内。 性成熟个体的体腔中充满近圆球形的具许多分散 卵黄和油滴的卵或有尾的精子。据统计,一条长 3─4cm 的雌虫可产卵达30000 万粒。 1.沙蚕的有性生殖 沙蚕科、裂虫科和矶沙 蚕科中的某些种类,于生殖前多发生形态上的变 化—生殖态,其中沙蚕科多数典型的海洋种还具 有特殊的群浮和婚舞的生殖现象。 生殖态(epitoky)是多数典型的海洋多毛类 中,于生殖前多发生形态上的变化的一种特有的生殖现象,是由无性或非生殖个体(atoke)

向浮游的有性或生殖个体(epitoke)转变过程,这种具有生殖态的虫体又称异沙蚕体 (heterouereis),在形体上均发生许多变化。主要表现在:眼变大,具晶体:围口触须变长 体中、后部疣足舌叶加宽扁:疣足背、腹须加长,雄性体前部疣足背须和腹须膨大:雄性除 体前部彭大外,体中、后部背须有锯齿状排列的乳突:所有刚毛逐步由游泳的奖状的刚毛所 取代:疣足肌显著拉长。上述种种变化有利于虫体有底栖转入暂时性的浮游,并使生殖细胞 不仅获得足够的营养,而且获得宽散的发育空间,使生殖产物易于排放。 多数沙蚕的精、卵成熟时经体壁的临时裂口排出体外,在海水中行体外受精。受精卵或 分散沉落粘附在它物上,如双齿围沙蚕、日本刺沙蚕等:或聚集于胶质物中形成卵块,如旗 须沙蚕、红角沙蚕Ceratonereis erythaeensis等。有少数种类如巨阔沙蚕Platgnereis eg阳lopS,雄虫常把尾部插入雌虫口内,被咬断后,精子穿过雌虫的咽进入雌虫体腔达到受 精的目的,进行体内受精。 2.沙蚕的发有受精卵多为黄卵,同多毛类的发有过程一样,一般需经过早期胚胎发有 (arly embryonic developmet)、担轮幼虫(ochophoraste、)后担轮幼虫(Metatro chophor stage),疣足幼虫(Nectochaeta stage)和刚节幼体(Setigger juvenile stage)各期成为成体。如日 本刺沙蚕各期形态有以下特征: (1)早期胚胎发有。卵子受精后,在卵膜内发有,约两个小时出现极体,四小时后行 第一次不等分裂,成为2细胞期,五小时后行第二次卵裂为四个不等大的分裂球。六小时后 行第三次明显的螺旋卵裂,所得八个分裂球,排成两层,每层四个,上层小,下层大。卵裂 继续进行,经无腔囊胚并以外包为主,内褶为辅进入原肠胚期,时为受精后一日。 (2)担轮幼虫。受精后两日,胚体出卵膜,为梨形,具口前纤毛轮、顶纤毛束、端纤 毛轮和端纤毛束。在口前纤毛轮前区具一对红色眼点。具一个分节的原始体腔、肠及原肾。 般发生在受精后2d。 (3)后担轮幼虫期。此期幼虫的标志是体节的出现。受精后4d,虫体二侧出现3对小 突起,为疣足刚毛原基,刚毛3束,但刚毛束伸出体外。疣足原基不能活动。口前纤毛轮的 中央出现多肌肉的咽原基,在口前纤毛轮与端纤毛轮之间具3个纤毛轮,属多纤毛轮幼虫 (polytrochal larva)。虫体具向光性,在培养皿中游动很快。 (4)疣足幼虫期。受精5五日后,刚毛伸出虫体外,此时疣足突起并逐渐分化成疣足 叶:顶纤毛束和端纤毛束消失,在虫体前端出现触手、触角和一圣触须突起,后端出现两个 肛突,它们皆先后变长分别发有形成触手、触角、一对触须和肛须,咽囊内出现大颚,消化 道逐渐分化为胃、肠等结构,并以肛门与体外相通。 (5)刚节幼体期受精后钓一个月,虫体第一对疣足前伸,刚毛脱落形成第二对触须, 该体节是形成围口节的一部分。此时疣足10对,无论在形态上和生态上都与成体相似,只 是性不成熟,故称刚节幼体
向浮游的有性或生殖个体(epitoke)转变过程,这种具有生殖态的虫体又称异沙蚕体 (heterouereis),在形体上均发生许多变化。主要表现在:眼变大,具晶体;围口触须变长; 体中、后部疣足舌叶加宽扁;疣足背、腹须加长,雌性体前部疣足背须和腹须膨大;雄性除 体前部彭大外,体中、后部背须有锯齿状排列的乳突;所有刚毛逐步由游泳的奖状的刚毛所 取代;疣足肌显著拉长。上述种种变化有利于虫体有底栖转入暂时性的浮游,并使生殖细胞 不仅获得足够的营养,而且获得宽敞的发育空间,使生殖产物易于排放。 多数沙蚕的精、卵成熟时经体壁的临时裂口排出体外,在海水中行体外受精。受精卵或 分散沉落粘附在它物上,如双齿围沙蚕、日本刺沙蚕等;或聚集于胶质物中形成卵块,如旗 须沙蚕、红角沙蚕 Ceratonereis erythaeensis 等。有少数种类如巨阔沙蚕 Platgnereis megalops,雄虫常把尾部插入雌虫口内,被咬断后,精子穿过雌虫的咽进入雌虫体腔达到受 精的目的,进行体内受精。 2.沙蚕的发育 受精卵多为黄卵,同多毛类的发育过程一样,一般需经过早期胚胎发育 (early embryonic development)、担轮幼虫(trochophora stage)、后担轮幼虫(Metatro chophora stage),疣足幼虫(Nectochaeta stage)和刚节幼体(Setigger juvenile stage)各期成为成体。如日 本刺沙蚕各期形态有以下特征: (1)早期胚胎发育。卵子受精后,在卵膜内发育,约两个小时出现极体,四小时后行 第一次不等分裂,成为2细胞期,五小时后行第二次卵裂为四个不等大的分裂球。六小时后 行第三次明显的螺旋卵裂,所得八个分裂球,排成两层,每层四个,上层小,下层大。卵裂 继续进行,经无腔囊胚并以外包为主,内褶为辅进入原肠胚期,时为受精后一日。 (2)担轮幼虫。受精后两日,胚体出卵膜,为梨形,具口前纤毛轮、顶纤毛束、端纤 毛轮和端纤毛束。在口前纤毛轮前区具一对红色眼点。具一个分节的原始体腔、肠及原肾。 一般发生在受精后 2d。 (3)后担轮幼虫期。此期幼虫的标志是体节的出现。受精后 4d,虫体二侧出现 3 对小 突起,为疣足刚毛原基,刚毛 3 束,但刚毛束伸出体外。疣足原基不能活动。口前纤毛轮的 中央出现多肌肉的咽原基,在口前纤毛轮与端纤毛轮之间具 3 个纤毛轮,属多纤毛轮幼虫 (polytrochal larva)。虫体具向光性,在培养皿中游动很快。 (4)疣足幼虫期。受精 5d 五日后,刚毛伸出虫体外,此时疣足突起并逐渐分化成疣足 叶;顶纤毛束和端纤毛束消失,在虫体前端出现触手、触角和一圣触须突起,后端出现两个 肛突,它们皆先后变长分别发育形成触手、触角、一对触须和肛须,咽囊内出现大颚,消化 道逐渐分化为胃、肠等结构,并以肛门与体外相通。 (5)刚节幼体期 受精后约一个月,虫体第一对疣足前伸,刚毛脱落形成第二对触须, 该体节是形成围口节的一部分。此时疣足10对,无论在形态上和生态上都与成体相似,只 是性不成熟,故称刚节幼体