
回 多肽链在超二级结构的基础上进一步折叠成紧密的近乎球状的结构,称为结构域 资 折桑子反映了一个蛋白质核心结构的拓扑结构模式,包含了蛋白质分子空间结物组 成的三个主要方面:即二级结构单元,二级结构单元的相对排列位置关系,以及多肽链 的走向。 折叠子主要有4种结构类型,全a蛋白、全B蛋白、B蛋白和+B蛋白。蛋白质 种折叠类型的定义为: 全蛋白一 一Q螺旋含量大于40%,B折叠含量小于5% 全B蛋白 -u螺旋含最小于5%,B折叠含量大于40% /B蛋白一 一心15%,伦15%。而且两种构象单元在肽链中是交替出现的。 ap蛋白 15%。两种构象单元分别聚集和分布在不同的区域 对只有单个结构域的蛋白。 结构域就是三维结构。对多结构域蛋白质,结构域与 构域之间以松散的肽段连接起来,组装成蛋白质的三级结构。 蛋白质的四级结构这个概念最早是由Bernal于1958年提出的,是指亚基与亚基之间通 过疏水相互作用,结合成为有序排列的空间结构。 习题和思考: 1、细胞学说的基本内容 2、说明细胞的分类 3、直核细的的基本结松 4 陈述蛋白质的生物学功能 试按你的理解对20种氨基酸进行分类 6、画图说明肽键的形成过程 何谓蛋白质一级结构,二级结构,超二级结构和四级结构? 课堂小结:1、要讲出与生物学其他课程的特别之处,主要是深度、新意 2、概括讲解即可 3、增加课堂互动, 特别是留给学生时间,让他们自己领悟
15 多肽链在超二级结构的基础上进一步折叠成紧密的近乎球状的结构,称为结构域。 折叠子反映了一个蛋白质核心结构的拓扑结构模式,包含了蛋白质分子空间结构组 成的三个主要方面:即二级结构单元,二级结构单元的相对排列位置关系,以及多肽链 的走向。 折叠子主要有 4 种结构类型,全 α 蛋白、全 β 蛋白、α/β 蛋白和 α+β 蛋白。蛋白质 4 种折叠类型的定义为: 全 α 蛋白——α 螺旋含量大于 40%,β 折叠含量小于 5% 全 β 蛋白——α 螺旋含量小于 5%,β 折叠含量大于 40% α/β 蛋白——α≥15%,β≥15%。而且两种构象单元在肽链中是交替出现的。 α+β 蛋白——α≥15%,β≥15%。两种构象单元分别聚集和分布在不同的区域。 对只有单个结构域的蛋白,结构域就是三维结构。对多结构域蛋白质,结构域与结 构域之间以松散的肽段连接起来,组装成蛋白质的三级结构。 蛋白质的四级结构这个概念最早是由 Bernal 于 1958 年提出的,是指亚基与亚基之间通 过疏水相互作用,结合成为有序排列的空间结构。 习题和思考: 1、 细胞学说的基本内容 2、 说明细胞的分类 3、 真核细胞的基本结构 4、 陈述蛋白质的生物学功能 5、 试按你的理解对 20 种氨基酸进行分类 6、 画图说明肽键的形成过程 何谓蛋白质一级结构,二级结构,超二级结构和四级结构? 课堂小结: 1、要讲出与生物学其他课程的特别之处,主要是深度、新意 2、概括讲解即可。 3、增加课堂互动,特别是留给学生时间,让他们自己领悟
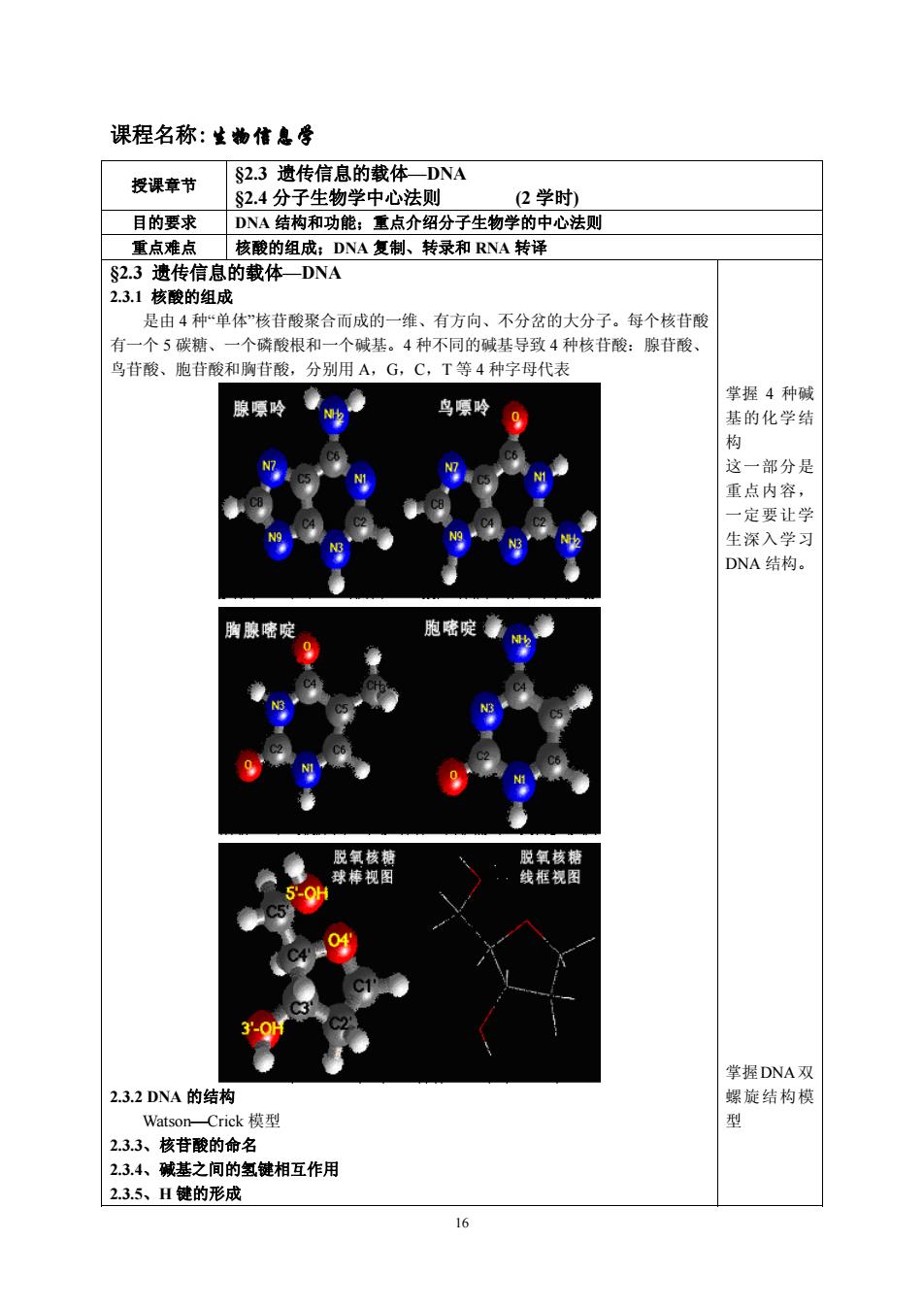
课程名称:生物信息辱 授课章节 §2.3遗传信息的载体一DNA S2.4分子生物学中心法则 (2学时) 目的要求 DNA结构和功能:重点介绍分子生物学的中心法则 重点难点核酸的组成:DNA复制、转录和RNA转译 S2.3遗传信息的载体DNA 231核酸的组成 是由4种“单体”核苷酸聚合而成的一维、有方向、不分岔的大分子。每个核苷酸 有一个5碳糖、一个磷酸根和一个碱基。4种不同的碱基导致4种核苷酸:腺苷酸、 鸟苷酸、胞苷酸和胸苷酸,分别用A,G,C,T等4种字母代表 鸟嘌吟 掌握4种碱 基的化学结 这一部分是 重点内容, 一定让学 生深入学习 DNA结构。 胞嘧啶章 脱氧核糖 脱氧核 线框视图 誉握DNA双 2.3.2DNA的结构 螺旋结构树 Watson-Crick模型 2.3.3、核苷酸的命名 2.3,4、碱基之间的氢键相互作用 23.5、H键的形成
16 课程名称:生物信息学 授课章节 §2.3 遗传信息的载体—DNA §2.4 分子生物学中心法则 (2 学时) 目的要求 DNA 结构和功能;重点介绍分子生物学的中心法则 重点难点 核酸的组成;DNA 复制、转录和 RNA 转译 §2.3 遗传信息的载体—DNA 2.3.1 核酸的组成 是由 4 种“单体”核苷酸聚合而成的一维、有方向、不分岔的大分子。每个核苷酸 有一个 5 碳糖、一个磷酸根和一个碱基。4 种不同的碱基导致 4 种核苷酸:腺苷酸、 鸟苷酸、胞苷酸和胸苷酸,分别用 A,G,C,T 等 4 种字母代表 2.3.2 DNA 的结构 WatsonCrick 模型 2.3.3、核苷酸的命名 2.3.4、碱基之间的氢键相互作用 2.3.5、H 键的形成 掌握 4 种碱 基的化学结 构 这一部分是 重点内容, 一定要让学 生深入学习 DNA 结构。 掌握DNA双 螺旋结构模 型
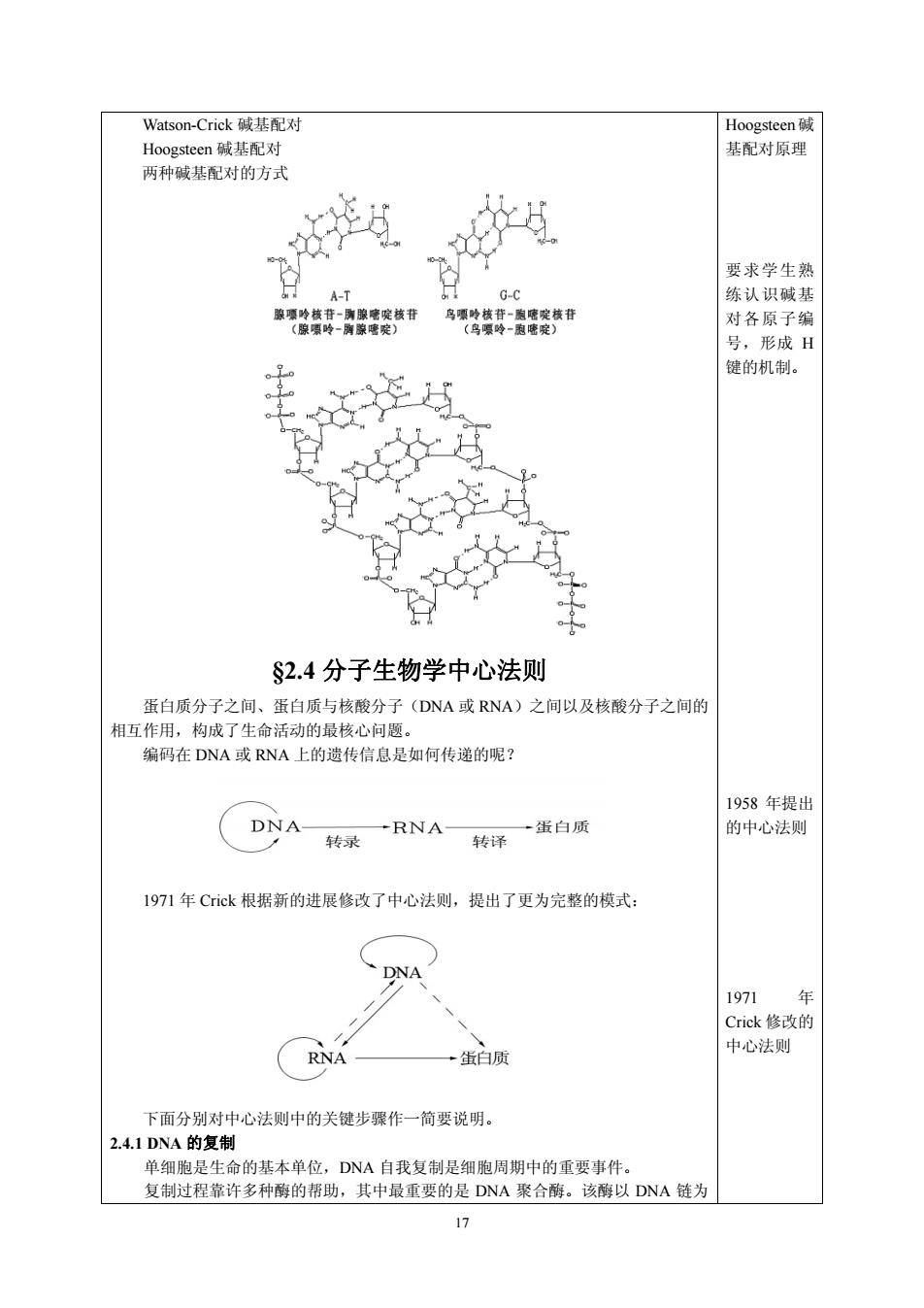
Watson-Crick碱基配对 Hoogsteen 基配对原理 要求学生朝 A-T G-c 练认识碱 改终琴宽族行 务修终哥:贵终技开 对各原子编 号,形成H 键的机制。 S2.4分子生物学中心法则 蛋白质分子之间、蛋白质与核酸分子(DNA或RNA)之间以及核酸分子之间的 相互作用,构成了生命活动的最核心问题。 编码在DNA或RNA上的遗传信息是如何传递的呢? 1958年提出 DNA 转录 RNA- 转译 蛋白质 的中心法 1971年Cick根据新的进展修改了中心法则,提出了更为完整的模式: 1971 年 Crick修改的 R 蛋白质 中心法则 下面分别对中心法则中的关键步骤作一简要说明。 2.4.1DNA的复制 单细胞是生命的基本单位,DNA自我复制是细胞周期中的重要事件。 复制过程靠许多种酶的帮助,其中最重要的是DNA聚合酶。该酶以DNA链为
17 Watson-Crick 碱基配对 Hoogsteen 碱基配对 两种碱基配对的方式 §2.4 分子生物学中心法则 蛋白质分子之间、蛋白质与核酸分子(DNA 或 RNA)之间以及核酸分子之间的 相互作用,构成了生命活动的最核心问题。 编码在 DNA 或 RNA 上的遗传信息是如何传递的呢? 1971 年 Crick 根据新的进展修改了中心法则,提出了更为完整的模式: 下面分别对中心法则中的关键步骤作一简要说明。 2.4.1 DNA 的复制 单细胞是生命的基本单位,DNA 自我复制是细胞周期中的重要事件。 复制过程靠许多种酶的帮助,其中最重要的是 DNA 聚合酶。该酶以 DNA 链为 Hoogsteen碱 基配对原理 要求学生熟 练认识碱基 对各原子编 号,形成 H 键的机制。 1958 年提出 的中心法则 1971 年 Crick 修改的 中心法则
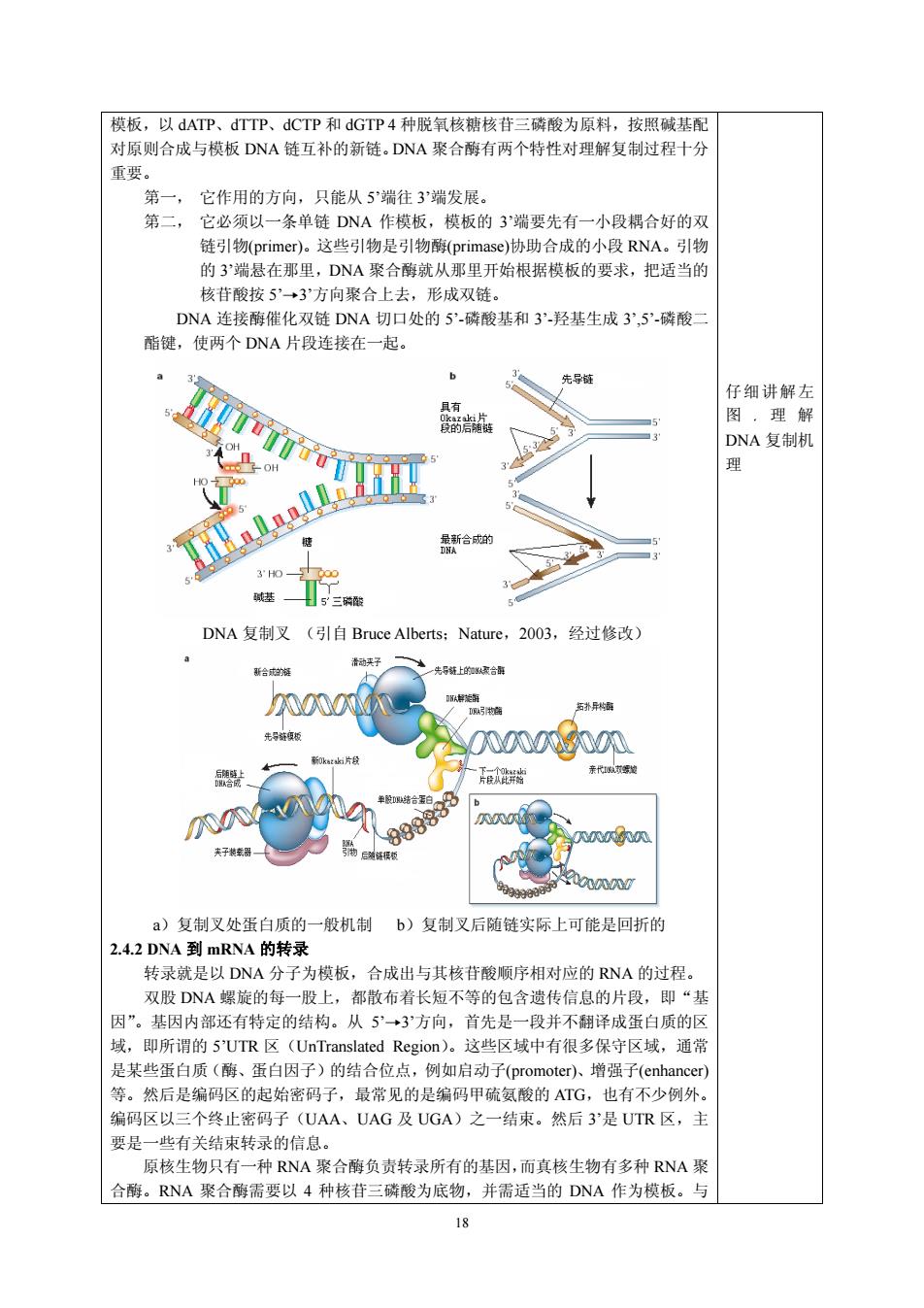
模板,以dATP、dTTP、dCTP和dGTP4种脱氧核糖核苷三磷酸为原料,按照碱基配 对原则合成与模板DNA链互补的新链.DNA聚合酶有两个特性对理解复制过程十分 重要 第一, 它作用的方向,只能从5端往3”端发展 第二, 它必须以一条单链DNA作模板,模板的3'端要先有一小段耦合好的双 链引物primer)。这些引物是引物(primase)协助合成的小段RNA。引物 的3端悬在那里,DNA聚合酶就从那里开始根据模板的要求,把话当的 核苷酸按5”→3”方向聚合上去,形成双链。 DNA连接酶催化双链DNA切口处的5-酸基和3羟基生成3,5-磷酸 酯键,使两个DNA片段连接在一起。 b 出导 仔细讲解左 图,理解 新合 基 5三时 DNA复制叉(引自Bruce Alberts::Nature.,2003,经过修改) 988 w a)复制叉处蛋白质的一般机制b)复制叉后随链实际上可能是回折的 2.42DNA到mRNA的转录 转录就是以DNA分子为模板,合成出与其核苷酸顺序相对应的RNA的过程 双股DNA螺旋的每一股上,都散布者长短不等的包含遗传信息的片段,即“去 因”。基因内部还有特定的结构。从5”→3方向,首先是一段并不翻译成蛋白质的区 域,即所谓的5'UTR区(UnTranslated Region)。这些区域中有很多保守区域,通常 是某些蛋白质(酶、蛋白因子)的结合位点,例如启动 、增强子心 等。然后是编码区的起始密码子, 最常见的是编码甲硫 AIG 也有不少例外 编码区以三个终止密码子(UAA、UAG及UGA)之一结束。然后3是UTR区,主 要是一些有关结束转录的信息。 原核生物只有一种RNA聚合酶负责转录所有的基因,而真核生物有多种RNA聚 合酶。RNA聚合酶需要以4种核苷三磷酸为底物,并需适当的DNA作为模板。与 18
18 模板,以 dATP、dTTP、dCTP 和 dGTP 4 种脱氧核糖核苷三磷酸为原料,按照碱基配 对原则合成与模板 DNA 链互补的新链。DNA 聚合酶有两个特性对理解复制过程十分 重要。 第一, 它作用的方向,只能从 5’端往 3’端发展。 第二, 它必须以一条单链 DNA 作模板,模板的 3’端要先有一小段耦合好的双 链引物(primer)。这些引物是引物酶(primase)协助合成的小段 RNA。引物 的 3’端悬在那里,DNA 聚合酶就从那里开始根据模板的要求,把适当的 核苷酸按 5’→3’方向聚合上去,形成双链。 DNA 连接酶催化双链 DNA 切口处的 5’-磷酸基和 3’-羟基生成 3’,5’-磷酸二 酯键,使两个 DNA 片段连接在一起。 DNA 复制叉 (引自 Bruce Alberts;Nature,2003,经过修改) a)复制叉处蛋白质的一般机制 b)复制叉后随链实际上可能是回折的 2.4.2 DNA 到 mRNA 的转录 转录就是以 DNA 分子为模板,合成出与其核苷酸顺序相对应的 RNA 的过程。 双股 DNA 螺旋的每一股上,都散布着长短不等的包含遗传信息的片段,即“基 因”。基因内部还有特定的结构。从 5’→3’方向,首先是一段并不翻译成蛋白质的区 域,即所谓的 5’UTR 区(UnTranslated Region)。这些区域中有很多保守区域,通常 是某些蛋白质(酶、蛋白因子)的结合位点,例如启动子(promoter)、增强子(enhancer) 等。然后是编码区的起始密码子,最常见的是编码甲硫氨酸的 ATG,也有不少例外。 编码区以三个终止密码子(UAA、UAG 及 UGA)之一结束。然后 3’是 UTR 区,主 要是一些有关结束转录的信息。 原核生物只有一种 RNA 聚合酶负责转录所有的基因,而真核生物有多种 RNA 聚 合酶。RNA 聚合酶需要以 4 种核苷三磷酸为底物,并需适当的 DNA 作为模板。与 仔细讲解左 图 , 理 解 DNA 复制机 理
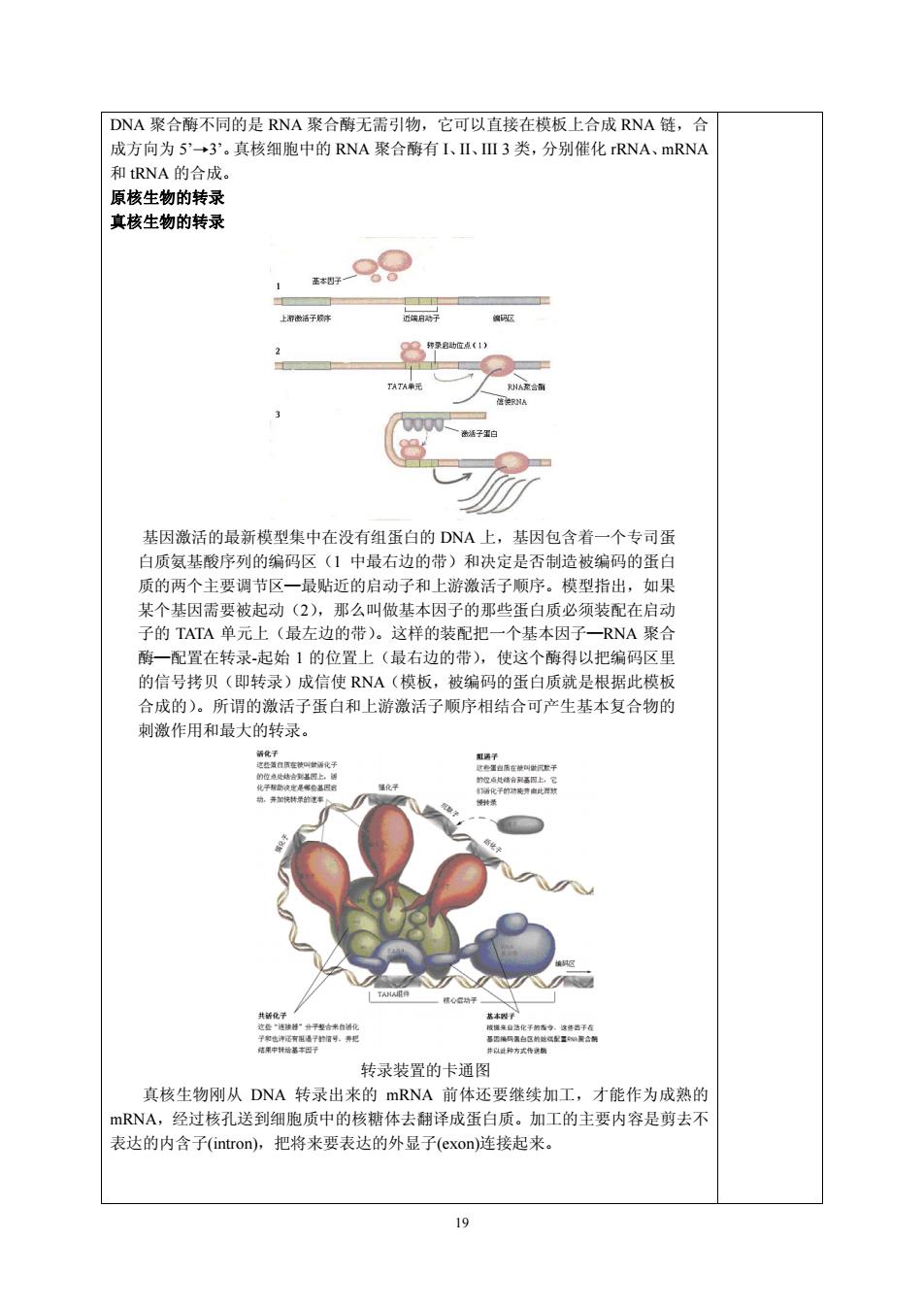
DNA聚合酶不同的是RNA聚合酶无需引物,它可以直接在模板上合成RNA链,合 成方向为5'→3'。真核细胞中的RNA聚合酶有I、Ⅲ3类,分别催化RNA、mRNA 和RNA的合成 原核生物的转录 真核生物的转录 99 上新 TATA 基因激活的最新模型集中在没有组蛋白的DNA上,基因包含者一个专司蛋 白质氨基酸序列的编码区(1中最右边的带 和决定是否制造被编码的蛋 质的两个主要调节区一最贴近的启动子和上游激活子顺序。模型指出,如果 某个基因需要被起动(2),那么叫做基本因子的那些蛋白质必须装配在启动 子的TATA单元上(最左边的带)。这样的装配把一个基本因子一RNA聚合 酶一配置在转录起始1的位置上(最右边的带).使这个酯得以把编码区里 的信号拷贝(即转录)成信使RNA(模板,被编码的蛋白质就是根据此模板 合成的)。所谓的激活子蛋白和上游激活子顺序相结合可产生基本复合物的 激作用和最大的转录。 转录装置的卡通图 真核生物刚从DNA转录出来的mRNA前体还要继续加工,才能作为成熟的 mRNA,经过核孔送到细胞质中的核糖体去翻译成蛋白质。加工的主要内容是剪去不 表达的内含子(intron),把将来要表达的外显子(exon)连接起来
19 DNA 聚合酶不同的是 RNA 聚合酶无需引物,它可以直接在模板上合成 RNA 链,合 成方向为 5’→3’。真核细胞中的 RNA 聚合酶有 I、II、III 3 类,分别催化 rRNA、mRNA 和 tRNA 的合成。 原核生物的转录 真核生物的转录 基因激活的最新模型集中在没有组蛋白的 DNA 上,基因包含着一个专司蛋 白质氨基酸序列的编码区(1 中最右边的带)和决定是否制造被编码的蛋白 质的两个主要调节区—最贴近的启动子和上游激活子顺序。模型指出,如果 某个基因需要被起动(2),那么叫做基本因子的那些蛋白质必须装配在启动 子的 TATA 单元上(最左边的带)。这样的装配把一个基本因子—RNA 聚合 酶—配置在转录-起始 1 的位置上(最右边的带),使这个酶得以把编码区里 的信号拷贝(即转录)成信使 RNA(模板,被编码的蛋白质就是根据此模板 合成的)。所谓的激活子蛋白和上游激活子顺序相结合可产生基本复合物的 刺激作用和最大的转录。 转录装置的卡通图 真核生物刚从 DNA 转录出来的 mRNA 前体还要继续加工,才能作为成熟的 mRNA,经过核孔送到细胞质中的核糖体去翻译成蛋白质。加工的主要内容是剪去不 表达的内含子(intron),把将来要表达的外显子(exon)连接起来