
第一节免疫应答的概念与过程 一、免疫应答的概念 抗原性物质进入机体后激发免疫细胞活化,分化和效应过程称之为免疫应答(immuner心sponse心),但对这一过程的认识是随着免疫学的发 展逐步深化和建立较为完整的概念。在免疫学发展的早期,人们发现抗原性物质进入体内后可在血液中检测出抗体,而对抗原是不产生应答 的,其指导学说为侧链学说和指令学说。 进入近代免疫学时朗,已知淋巴细跑可能是抗体产生细胞,但尚无直接证明,实验认为抗原只对抗体产生细胞起选择激发作用,并且认为 抗体只是由单一淋巴细胞克隆完成的,彼此无关联。在这一时期已证明除存在体液免疫外,还存在有细息免疫对自己抗原提出了自身耐受 的概念,在打破自身耐受的条件下可以发生自身免疫现象,并证明在自身免疫性疾病患者血清中有自身抗体存在。这一时期的指导学说是由 Bumc提出的细胞系或克降选择学说. 进入现代免疫学时期,已证明在高等动物和人体内存在有结构复杂的免疫系统,是由免疫器直、兔疫细胞和免疫分子组成的。同时也证明 了免疫应答是由多细胞系完成的。并认为它们之间存在着相互协同的和相互制约的关系,在正常免疫生理条件下,它们处于动态平衡,借以维 持机体的免疫稳定状态抗原进入可激发免疫系统打破了这种平衡,从而诱发了免疫应答,再建立新的平衡状态。在此时棚还证明了自身耐受 是由多种机制形成的,存在着生理性自身识别”与病理性自身识其指导学说是由 担出的学说 二、免疫应答的过程 现代免疫学的发展,为了解免疫应答过程提供了细胞学基础。虽然免疫应答过程的效应表现主要是以B细胞介导的体液免疫和以T细胞介 和体 的已明 ,这二神免应答的产生都是 由多细跑系完成的,即由单核吞噬细胞 元的 细胞系的行为,而是 多细胞系相 作用的复杂行为.。这一-过程包活:①免陵细胞对抗原分子的识别过程,即抗原分 子与免疫细胞的相互作用:②免疫细胞的活化和分化过程,即免疫细胞间的相互作用效应细购和效应分子的排异作用。(表111) 表11免疫应答过程 的作甲 生与传递 抗原的摄取处理和加工抗系的呈抗原的识 引起组织的损作用(炎症)】 细孢增殖与分化 免陵应答的调节作用 生物活性介质的合成与释放 抗分 细 免疫增系统 补体分子 抗 用子 细因子的产生与程 KNK细跑 效应T细胞: 细胞的产生 细胞系 宿主 对免疫应答过程的了解,就是安对上述过程进行深入的深讨,现已主证明免疫细狗间的相互作用,既表现相互协同又表现相互拮抗, 细胞间的相互作用是由其分论的介质充成的。近年来的实验又证明,免疫细胞在其相互作用中,可表现为具有组织相容性基因的限制性,即所 调MHC限制性,表明免痘细胞的活动是受遗传控制的。上述发现,既揭示了免疫应答过程的复杂性,又反映了它是严密控制和精细的调节过 程,这对保持机体自身免疫稳定性是十分重要的。因此,目前对免疫应答机制的研究,已由细胞水平。分子水平进入了基因水平,虽然如此, 对这一复杂过程的认识是极其有限的,众多的问题还有待解决 1,抗原识别阶段此阶段可包括抗原的摄取,处理和加工,抗原的呈递和对抗原的识别,分别由M、T和B细胞完成 免疫细抱的活化和分化阶段此阶段可包括抗原识别细跑膜受体的交联、膜信号产生与传递、细抱蜡殖与分化以及生物活性介质的合成与 释故,主要由和B细狗完成。 ,免疫应答的效应阶段此阶段主要包括效应分子(体液免疫)和效应细胞(细胞免疫)对非已细跑或分子的清除作用,即所调排异效应 及其对免疫应答的调节作用。此阶段除抗体和效应T细胞参予外,即非特异免疫细胞和分子参加才能完成排异和免疫调节作用。 第二节抗体产生的细胞学基础 一、抗体产生的一般规律 当第一次用适量抗原给动物免疫,需经一定伏期才能在血液中出现抗体,含量低,且维特时间短,很快下隆,称这种现象为初次免疫应 答。若在抗体下降期再次给以相同抗原免疫时,则发现抗体出现的潜伏期较初次应答明显缩短,抗体含量也随之上升,而且维持时间长,称这
第一节 免疫应答的概念与过程 一、免疫应答的概念 抗原性物质进入机体后激发免疫细胞活化,分化和效应过程称之为免疫应答(immuneresponse)。但对这一过程的认识是随着免疫学的发 展逐步深化和建立较为完整的概念。在免疫学发展的早期,人们发现抗原性物质进入体内后可在血液中检测出抗体,而对抗原是不产生应答 的,其指导学说为侧链学说和指令学说。 进入近代免疫学时期,已知淋巴细胞可能是抗体产生细胞,但尚无直接证明,实验认为抗原只对抗体产生细胞起选择激发作用,并且认为 抗体只是由单一淋巴细胞克隆完成的,彼此无关联。在这一时期已证明除存在体液免疫外,还存在有细胞免疫。对“自己”抗原提出了自身耐受 的概念,在打破自身耐受的条件下可以发生自身免疫现象,并证明在自身免疫性疾病患者血清中有自身抗体存在。这一时期的指导学说是由 Burnet提出的细胞系或克隆选择学说。 进入现代免疫学时期,已证明在高等动物和人体内存在有结构复杂的免疫系统,是由免疫器官、免疫细胞和免疫分子组成的。同时也证明 了免疫应答是由多细胞系完成的。并认为它们之间存在着相互协同的和相互制约的关系,在正常免疫生理条件下,它们处于动态平衡,借以维 持机体的免疫稳定状态。抗原进入可激发免疫系统打破了这种平衡,从而诱发了免疫应答,再建立新的平衡状态。在此时期还证明了自身耐受 是由多种机制形成的,存在着生理性“自身识别”与病理性“自身识别”。其指导学说是由Jerne提出的免疫网络学说。 二、免疫应答的过程 现代免疫学的发展,为了解免疫应答过程提供了细胞学基础。虽然免疫应答过程的效应表现,主要是以B细胞介导的体液免疫和以T细胞介 导的细胞免疫。但体内和体外的实验已证明,这二种免疫应答的产生都是由多细胞系完成的,即由单核吞噬细胞系、T细胞和B细胞来完成的。 所以免疫应答过程不是单一细胞系的行为,而是多细胞系相互作用的复杂行为。这一过程包括:①免疫细胞对抗原分子的识别过程,即抗原分 子与免疫细胞的相互作用;②免疫细胞的活化和分化过程,即免疫细胞间的相互作用;③效应细胞和效应分子的排异作用。(表11-1)。 表11-1 免疫应答过程 识别 活化与分化 效应 抗原的摄取、处理和加工抗原的呈递抗原的识别 膜受体的交联 膜信号的产生与传递 细胞增殖与分化 生物活性介质的合成与释放 对靶细胞或分子的排异作用 引起组织的损作用(炎症) 免疫应答的调节作用 抗原→ Mφ T B T与B细胞的境殖与分化 抗体的产生与释放 细胞因子的产生与释放 效应T细胞的产生 免疫记忆细胞的产生 抗体分子 效应T细胞 免疫增强系统 补体分子 细胞因子 K/NK细胞 Mφ粒细胞系 肥大细胞 红细胞 血小板 排异 或排已 免疫保护 免疫病理 抗感染 抗肿 自身 变态 移植 移植 宿主 对免疫应答过程的了解,就是要对上述过程进行深入的探讨。现已主证明免疫细胞间的相互作用,既表现相互协同又表现相互拮抗,同时 细胞间的相互作用是由其分泌的介质完成的。近年来的实验又证明,免疫细胞在其相互作用中,可表现为具有组织相容性基因的限制性,即所 谓MHC限制性,表明免疫细胞的活动是受遗传控制的。上述发现,既揭示了免疫应答过程的复杂性,又反映了它是严密控制和精细的调节过 程,这对保持机体自身免疫稳定性是十分重要的。因此,目前对免疫应答机制的研究,已由细胞水平、分子水平进入了基因水平,虽然如此, 对这一复杂过程的认识是极其有限的,众多的问题还有待解决。 1.抗原识别阶段此阶段可包括抗原的摄取、处理和加工,抗原的呈递和对抗原的识别,分别由Mφ、T和B细胞完成。 2.免疫细胞的活化和分化阶段此阶段可包括抗原识别细胞膜受体的交联、膜信号产生与传递、细胞增殖与分化以及生物活性介质的合成与 释放,主要由T和B细胞完成。 3.免疫应答的效应阶段此阶段主要包括效应分子(体液免疫)和效应细胞(细胞免疫)对非已细胞或分子的清除作用,即所谓排异效应, 及其对免疫应答的调节作用。此阶段除抗体和效应T细胞参予外,即非特异免疫细胞和分子参加才能完成排异和免疫调节作用。 第二节 抗体产生的细胞学基础 一、抗体产生的一般规律 当第一次用适量抗原给动物免疫,需经一定潜伏期才能在血液中出现抗体,含量低,且维持时间短,很快下降,称这种现象为初次免疫应 答。若在抗体下降期再次给以相同抗原免疫时,则发现抗体出现的潜伏期较初次应答明显缩短,抗体含量也随之上升,而且维持时间长,称这

种现象为现次免疫应答或回忆应答。由于对抗体分子结构研究的进展,发现初次应答产生的抗体主要是M分子,对抗原结合力低,为低亲和 性抗体。而再次应答则主要为1gG分子,且为高亲和性抗体,TD抗原可引起再次应答,而T抗原只能引起切应答。对切次和再次应答现象机制 的研究,对抗体特异性、多样性、免疫记忆以及对自身抗原而受性机制等问题的研究,都必须以抗体生成的细跑学为基础(图111,表1山 2) 图11,1初次及再次免疫应答 表112初次与再次免夜应答特性 二、抗体产生的细胞学基甜 抗体产生是由多细胞完成的,M©等在60年代,首先证明了淋巴细胞是不均一的细群,他用早期摘除鸡的胸腺和法氏囊的方法证明了 有一 ,后者与抗体形成有关(表113)。 表113新生期滴除胸腺及法氏囊对免疫 能的影(鸡 抗体产生 法氏囊 13200 + +阳性反应:·阴性反应 给经X线照射小鼠移入同系骨菌细的(B细胞来源)和胸腺细跑(T细胸来源),然后用羊红细胞免疫,结果证明只有同时移入 两种细胞才能产生抗体,因此证明了抗体产生需要T和B细胞共同参予。 inanue等在70年代又证明了巨噬细胞在抗体形成中的重要作用。他们应用纯化细胞的体外培养技术研究这一问题。根暑小鼠细孢对玻璃面 或塑料面的粘附性,可将璃细胞分为二种,其一为有粘附性细抱属巨堂细跑(M),另一种为非粘附性细胞属淋巴细抱,包括T和B细抱,当 将这二种细胞分别与羊红细孢(抗原)在体外培养时,皆不能产生抗体,只有在二种细胞混合培养时才能产生抗体,自此证明了M也参予抗 体的产生(表114,5) 表114T和B细胞在抗体产生中的作用 抗体产生 表1 在抗体产生中的作用 抗体产 细 红细 非粘附细胞 表11-6促进B细胞增殖和分化的细胞因子 作用 L-4 进细作 IL-5 活化动 L-6 促进B细胞分化 IL-2 与L-1协同
种现象为现次免疫应答或回忆应答。由于对抗体分子结构研究的进展,发现初次应答产生的抗体主要是IgM分子,对抗原结合力低,为低亲和 性抗体。而再次应答则主要为IgG分子,且为高亲和性抗体。TD抗原可引起再次应答,而TI抗原只能引起初应答。对初次和再次应答现象机制 的研究,对抗体特异性、多样性、免疫记忆以及对自身抗原而受性机制等问题的研究,都必须以抗体生成的细胞学为基础(图11-1,表11- 2)。 图11-1初次及再次免疫应答 表11-2 初次与再次免疫应答特性 特性 初次 再次 抗原呈递 非B细胞 B细胞 抗原浓度 高 低 抗体产生 延迟相 5~10天 2~5天 Ig类别 主要为IgM IgG、IgA等 亲和力 低 高 无关抗体 多 少 二、抗体产生的细胞学基础 抗体产生是由多细胞完成的,Miller等在60年代,首先证明了淋巴细胞是不均一的细胞群。他用早期摘除鸡的胸腺和法氏囊的方法证明了 有二类不同的的淋巴细胞,即T和B细胞。前者与细胞免疫有关,后者与抗体形成有关(表11-3)。 表11-3 新生期摘除胸腺及法氏囊对免疫功能的影响(鸡) 全身X-线照射 周围血淋巴细胞数 Ig浓度 抗体产生 移植物排斥反应 未身X-线照射 148 000 ++ +++ ++ 胸腺摘除 9 000 ++ + - 法氏囊摘除 13 200 - - + +阳性反应;-阴性反应 Claman 给经X-线照射小鼠移入同系骨髓细胞(B细胞来源)和胸腺细胞(T细胞来源),然后用羊红细胞免疫,结果证明只有同时移入 两种细胞才能产生抗体。因此证明了抗体产生需要T和B细胞共同参予。 Unanue等在70年代又证明了巨噬细胞在抗体形成中的重要作用。他们应用纯化细胞的体外培养技术研究这一问题。根据小鼠细胞对玻璃面 或塑料面的粘附性,可将脾细胞分为二种,其一为有粘附性细胞属巨噬细胞(Mφ),另一种为非粘附性细胞属淋巴细胞,包括T和B细胞。当 将这二种细胞分别与羊红细胞(抗原)在体外培养时,皆不能产生抗体,只有在二种细胞混合培养时才能产生抗体,自此证明了Mφ也参予抗 体的产生(表11-4,5)。 表11-4 T和B细胞在抗体产生中的作用 X-线照射鼠入的细胞 抗体产生 脾细胞(含有T和B) ++ 胸腺细胞(T细胞) ± 骨髓细胞(B细胞) + 胸腺细胞+骨髓细胞 +++ 表11-5 Mφ在抗体产生中的作用 体外培养细胞 抗体产生 粘附细胞+羊红细胞 非粘附细胞+羊红细胞 粘附细胞 + +羊红细胞 非粘附细胞 -- +++ 表11-6 促进B细胞增殖和分化的细胞因子 名称 作用 IL-1 IL-4 IL-5 IL-6 IL-2 促进B细胞活化与增殖 促进活化B细胞增殖 促进B细胞分化 与IL-1协同

在同一时期Gershon等又证明了抑制性T细胞(TS)的存在,对免疫应答起抑制作用.因此TH和TS可视为免疫调节细胞,而TC和TD可视 为细胞免疫的效应细胞。通过上述研究,证明了抗体产生需要三种细胞参予,即单核吞噬细胞系。T细跑系和B细胞系。从而否定了过去认为抗 体产生是由单一淋巴细胞克隆产生的观点。因此抗体产生不只是涉及抗原与免疫细跑间的相互作用,即对抗原的识别和抗原的激发作用,同时 也涉及免疫细胞间的相互作用,,即免疫细陶活化,增殖与分化过程。这二个过程是紧密交织在一起的,为此必须进一步探讨在免疫应答过程 中,三种细胞各自发挥什么作用?以及它们之间的相互作用又是怎样进行的 三、免疫细胞在抗体生成中的作用 上述三类细跑都参予抗体生成过程,但各自发挥的作用不同。现已证明M抗原处理和呈递细抱,无特异识别抗原的功能。T细胞系主要是 TH和TS,它们对免疫应答有调节功能,所以是免疫调节细胞,有特异识抗原的功能。B细跑系既具有呈递抗原的作用又是产生抗体的细跑。 也且有特异识别抗原的功能 一}的作用 M在免疫应答的全过程都发挥重要作用,在抗原识别过程中,即在免疫应答的诱导期,它表现为具有摄取、处理加工、存贮和呈递抗原的 作用。它活化后还能分泌多种细抱因子,其合成和分论的L1有促进T和B细胞的活化作用。因此,不能认为M只是机械的将抗原决定簇呈递 给淋巴细胞,它还具有调节淋巴细跑功能的作用。 抗性物质讲入体内后,必须先经口取。加工处理,然后才号 多种受体分子,但无抗原识别受体,它主要是以吞磁、吞饮和被动吸附等方式捕捉抗原,可摄取任何抗原性特质,所以是非特异性的摄取抗 性物压 摄入的抗原大分子,可在细内被降解为许多小肽片段。其 一些免疫原性决定簇可与细跑内的自己MHC类分子相结合,然后运送至细 去面形成E 饰的自身复合物 M将这种复合物分子 抗原X+自己 识别这种复合物的抗原识别受体,必须是既能 ()淋巴细的作用 互作用 MHC限制性的 由来 淋巴细孢具有抗原识 抗原决定簇,所以这种识别是有特异性的。B年 胞表面抗原识别受 可识别天然 白质抗 餐体分子,即TCR 加工处理的序列决定肽片段(即T决定簇)和白己MHC分子 四、细胞因子在抗体产生中的作用 细跑因子(cytokine)在抗体产生应答过程中的作用有下述特点 1,细胞因子的作用既无抗原特异性也无MHC限制性当TH细跑受刺激活化后,它所分论的细泡因子就可作用于任何抗原特异性的B细胞和 任何MHC单倍型(haplotype)的B细跑. 2.在B细胞产生免疫应答的不同时期有不同的细胞因子起作用即B细胞的增殖期与分化期,或g的分论期可有不同的细胞因子在起作用 (表16)。此外,不同的细胞因子间的组合有的起结抗作用,有的起协同作用。 3.细跑因子还可作用于旁路(bystander)B细胞使之活化这些B细胞对抗体应答的抗原没有特异性(非抗原特异性B细胞),它们存在于 抗原刺激的特异B细胞周围,在抗原活化特异B细胞时,因产生细胞因子而被活化并产生非特异抗体。 4.在刺激B细陶塔殖分化的细跑因子中,有些是来源于巨噬细息或其他非T细胞所产生(如粒细胞。肥大细胞等), 第三节抗体形成过程中免疫细胞间的相互作用 在抗体生成过程中,对TD抗原的应答已证明是由M。,T系细胞和B系细胞相互作用的结果,那么M仰直接呈递抗原的对象是T细孢还是B细 胸?或者二者都是?就现有资料分析,它们可能存在着M0与T的相互作用,T与B的相互作用以及Mφ与B细胞之间的相互作用。 一、巨噬细胞与TH细胞之间的相互作用 TH细胞必须由静止状态的TH转变活化为H才能错助B细施产生抗体,关于TH细胞活化的机制,根蜗现有实验资料提出一个双信号假说, 近年的实验证明在抗原呈递细胞(APC)表面至少有二种分子与TH细胞的活化相关。 一种是抗原是浦分子,它是由C分子组成,它司 与外源性或内源性抗原欧片段结合,然后运送至细胞表面并呈递给 细胞,通过TCR/CE3刺激产生第1活化信号.另 子即所谓协同刺激分 子(costimulating 它导中 一组粘附分 组成,它不仅能 与T细胞的直接接触 传速的功能, 组分子可与T细上的协网刺激分子受体( MR) 结合 激其产生协同 信号,即所调装
在同一时期Gershon等又证明了抑制性T细胞(TS)的存在,对免疫应答起抑制作用。因此TH和TS可视为免疫调节细胞,而TC和TD可视 为细胞免疫的效应细胞。通过上述研究,证明了抗体产生需要三种细胞参予,即单核吞噬细胞系、T细胞系和B细胞系。从而否定了过去认为抗 体产生是由单一淋巴细胞克隆产生的观点。因此抗体产生不只是涉及抗原与免疫细胞间的相互作用,即对抗原的识别和抗原的激发作用,同时 也涉及免疫细胞间的相互作用,,即免疫细胞活化,增殖与分化过程。这二个过程是紧密交织在一起的,为此必须进一步探讨在免疫应答过程 中,三种细胞各自发挥什么作用?以及它们之间的相互作用又是怎样进行的。 三、免疫细胞在抗体生成中的作用 上述三类细胞都参予抗体生成过程,但各自发挥的作用不同。现已证明Mφ抗原处理和呈递细胞,无特异识别抗原的功能。T细胞系主要是 TH和TS,它们对免疫应答有调节功能,所以是免疫调节细胞,有特异识别抗原的功能。B细胞系既具有呈递抗原的作用又是产生抗体的细胞, 也具有特异识别抗原的功能。 (一)Mφ的作用 Mφ在免疫应答的全过程都发挥重要作用,在抗原识别过程中,即在免疫应答的诱导期,它表现为具有摄取、处理加工、存贮和呈递抗原的 作用。它活化后还能分泌多种细胞因子,其合成和分泌的IL-1有促进T和B细胞的活化作用。因此,不能认为Mφ只是机械的将抗原决定簇呈递 给淋巴细胞,它还具有调节淋巴细胞功能的作用。 抗原性物质进入体内后,必须先经Mφ摄取、加工处理,然后才能呈递给淋巴细胞。Mφ是有吞噬细胞功能的细胞,已证明在其细胞表面有 多种受体分子,但无抗原识别受体。它主要是以吞噬、吞饮和被动吸附等方式捕捉抗原,可摄取任何抗原性特质,所以是非特异性的摄取抗原 性物质。 摄入的抗原大分子,可在细胞内被降解为许多小肽片段。其中一些免疫原性决定簇可与细胞内的自己MHCⅡ类分子相结合,然后运送至细 胞膜表面,形成所谓修饰的自身复合物分子(即异种抗原X+自己MHCⅡ类分子)此即Mφ对抗原的处理和加工过程。 Mφ将这种复合物分子,呈递给有抗原识别功能的淋巴细胞,才能激发免疫应答。不难看出,识别这种复合物的抗原识别受体,必须是既能 识别异种抗原X,又能同时识别自已MHC分子。这就是免疫细胞间相互作用的MHC限制性的由来。 (二)淋巴细胞的作用 淋巴细胞具有抗原识别受体,所以T和B细胞都是抗原识别细胞。每一细胞克隆可识别一种抗原决定簇,所以这种识别是有特异性的。B细 胞表面抗原识别受体是膜Ig分子,它可识别天然蛋白质抗原分子表面的构像抗原决定簇(即B决定簇),在识别抗原时无MHc 限制性。而T细 胞抗原识别受体为异二聚体分子,即TCRαβ,它能同时识别经加工处理的序列决定簇肽片段(即T决定簇)和自己MHC分子,所以有MHC限制 性。 四、细胞因子在抗体产生中的作用 细胞因子(cytokines)在抗体产生应答过程中的作用有下述特点: 1.细胞因子的作用既无抗原特异性也无MHC限制性当TH细胞受刺激活化后,它所分泌的细胞因子就可作用于任何抗原特异性的B细胞和 任何MHC单倍型(haplotype)的B细胞。 2.在B细胞产生免疫应答的不同时期有不同的细胞因子起作用即B细胞的增殖期与分化期,或Ig的分泌期可有不同的细胞因子在起作用 (表11-6)。此外,不同的细胞因子间的组合有的起拮抗作用,有的起协同作用。 3.细胞因子还可作用于旁路(bystander)B细胞使之活化这些B细胞对抗体应答的抗原没有特异性(非抗原特异性B细胞)。它们存在于 抗原刺激的特异B细胞周围,在抗原活化特异B细胞时,因产生细胞因子而被活化并产生非特异抗体。 4.在刺激B细胞增殖分化的细胞因子中,有些是来源于巨噬细胞或其他非T细胞所产生(如粒细胞、肥大细胞等)。 第三节 抗体形成过程中免疫细胞间的相互作用 在抗体生成过程中,对TD抗原的应答已证明是由Mφ、T系细胞和B系细胞相互作用的结果,那么Mφ直接呈递抗原的对象是T细胞还是B细 胞?或者二者都是?就现有资料分析,它们可能存在着Mφ与T的相互作用,T与B的相互作用以及Mφ与B细胞之间的相互作用。 一、巨噬细胞与TH细胞之间的相互作用 TH细胞必须由静止状态的TH转变活化为TH才能辅助B细胞产生抗体。关于TH细胞活化的机制,根据现有实验资料提出一个双信号假说。 近年的实验证明在抗原呈递细胞(APC)表面至少有二种分子与TH细胞的活化相关。一种是抗原呈递分子,它是由MHC分子组成,它可 与外源性或内源性抗原肽片段结合,然后运送至细胞表面并呈递给T细胞,通过TCR/CE3刺激产生第1活化信号。另一种分子即所谓协同刺激分 子(costimulating molecules,CM),它是由一组粘附分子组成,它不仅能促进APC与T细胞的直接接触,而且也具有诱导信号传递的功能。这 组分子可与T细胞上的协同刺激分子受体(costimulatory moleculesrecptor,CMR)结合,刺激其产生协同激信号,即所谓第2信号
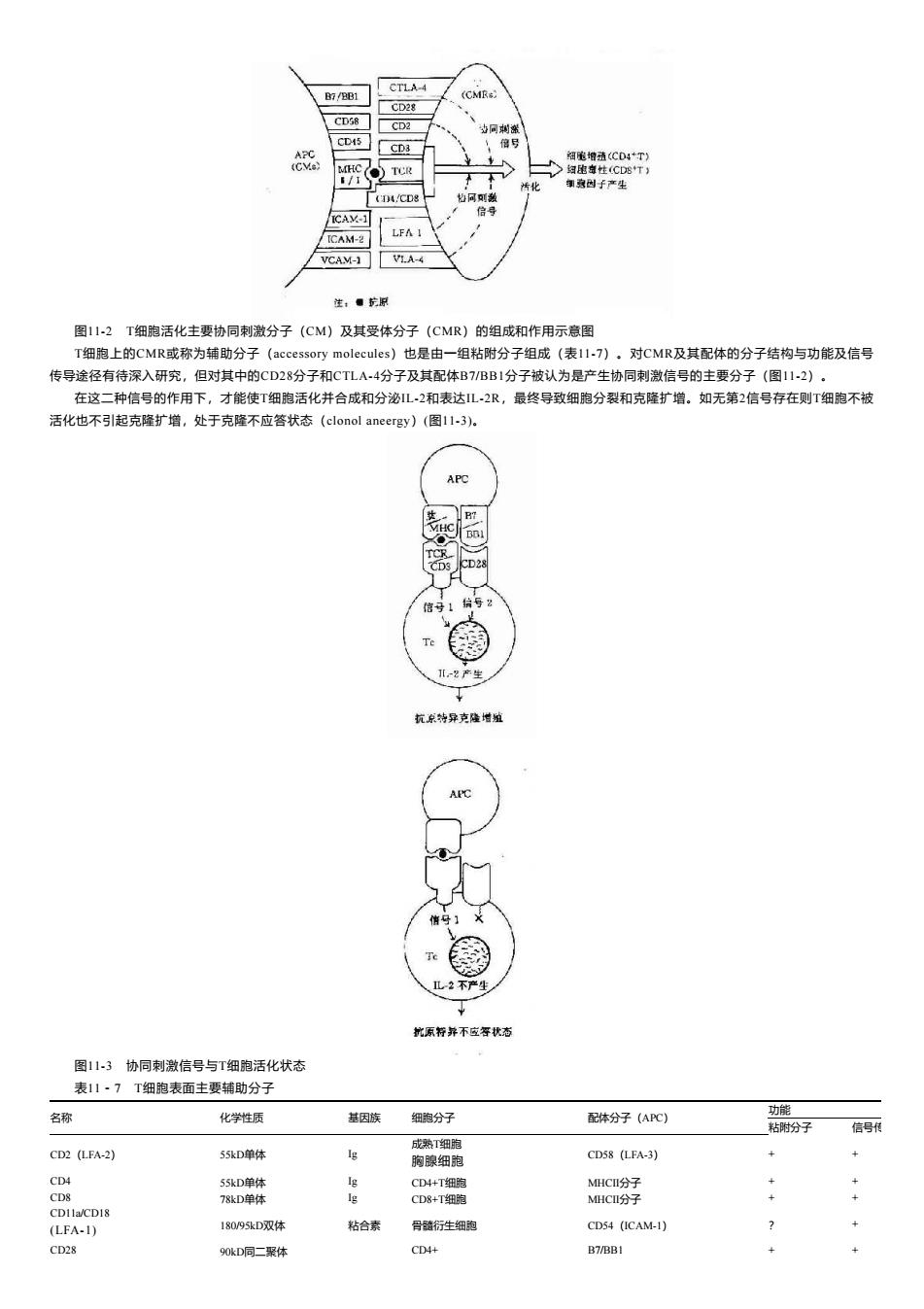
CD第 > VCAM-1 VLA-4 生,。发原 图1-2 T细胞活化主要协同刺激分子(CM)及其受体分子(CMR)的组成 作用示意图 「细胞上的 MR或称为辅助分子(accessory molecules)也是由 一组粘附分子组成(表117),对CMR及其配体的分子结构与功能及信号 传导途径有待深入研究 但对其中的CD28分子和CTLA4分子及其配体B7BB1分子被认为是产生协同刺激信号的主要分子(112)· 在这二种信号的作用下,才能使T细胞活化并合成和分泌L2和表达L2R,最终导致细跑分裂和克隆扩增。如无第2信号存在则T细胞不被 活化也不引起克隆扩增,处于克隆不应答状态(clonol aneergy)(图11-3), Te 优原并不空状 图11.3协同刺激信号与T细跑活化状态 表11·7T细胞表面主要辅助分子 名称 化学性质 族 细分子 信号 CD2 (LFA-2) 55kD单付 g 细 CD58 (LFA-3) 1809D双体 粘合泰 骨酷行生细的 CD54 (ICAM-1) 90D同二聚
图11-2 T细胞活化主要协同刺激分子(CM)及其受体分子(CMR)的组成和作用示意图 T细胞上的CMR或称为辅助分子(accessory molecules)也是由一组粘附分子组成(表11-7)。对CMR及其配体的分子结构与功能及信号 传导途径有待深入研究,但对其中的CD28分子和CTLA-4分子及其配体B7/BB1分子被认为是产生协同刺激信号的主要分子(图11-2)。 在这二种信号的作用下,才能使T细胞活化并合成和分泌IL-2和表达IL-2R,最终导致细胞分裂和克隆扩增。如无第2信号存在则T细胞不被 活化也不引起克隆扩增,处于克隆不应答状态(clonol aneergy)(图11-3)。 图11-3 协同刺激信号与T细胞活化状态 表11-7 T细胞表面主要辅助分子 名称 化学性质 基因族 细胞分子 配体分子(APC) 功能 粘附分子 信号传 CD2(LFA-2) 55kD单体 Ig 成熟T细胞 胸腺细胞 CD58(LFA-3) + + CD4 55kD单体 Ig CD4+T细胞 MHCⅡ分子 + + CD8 78kD单体 Ig CD8+T细胞 MHCⅡ分子 + + CD11a/CD18 (LFA-1) 180/95kD双体 粘合素 骨髓衍生细胞 CD54(ICAM-1) ? + CD28 90kD同二聚体 CD4+ B7/BB1 + +
CD8+T(50%】 .0 异二聚体 粘合 细外兰质VCAM.1 P3%CD40) CD4+细的 CD40 阻断或给予第2信号,可以人为调节免疫应答使之增强或或抑制,对免疫治疗提供了新的手段。如阻断第2信号的产生可使T细胞处于免疫 耐受状态,降低机体的免疫应答,这对防止移植排斥的发生和对超敏性疾病以及自身免疫性疾病的治疗是有利的。如将B7基因导入一定的肿粽 细胞则可增强机体的抗肿密免疫应答,目前这方面的实验研究已有较多的报道(图114)。 不花化 香议电波答, 冲爱免疫 基腾7Cc 密 图114协同刺激信号与免疫应答调 二、TH细胞与B细胞之间的相互作用 (一)B细胞对T细跑依赖抗原的应答 般情况下当大量抗原进入未经免疫的机体后,诱发初次免疫应答时其抗原呈递细胞多由巨噬细胞完成。经M活化TH细跑后再由活化的 TH细胞铺助B细跑产生抗体和形成记忆B细胞。但当再次免疫应答发生时,抗原呈递细跑则主要由已扩增的B细胞克隆承担。由于其膜g受体亲 和力增高故对少量抗原也能摄取,故可取代巨噬细胞的抗原呈递作用。 过去曾认为B细胞与TH细胞间的相互作用是通过抗原的桥联作用使之发生接触,即B细胞的抗原受体可识别抗原分子的半抗原部分,而 细胞的抗原受体可识别抗原分子的线体部分。但近年的实验证明B细胞与M0相同,也是通过抗原呈递作用与TH细胞相互作用。即B细胞可通过 其膜!E受体蛋白质抗原,即T依赖性抗原结合,经内化作用将抗原摄入,然后经过加工处理,使T细胞决定簇与MHC川类分子结合,形成MHC 肽分子复合物并运送至B细胞表面,将其呈递给TH细胞,所以B细抱与TH细抱间的相互作用也是MHC限制性的。 B细胞和TH细胞通过其粘附分子及抗原呈递作用可彼此直接接触,并能相互诱导使之活化。活化B细跑在TH细胞的辅助下,B细抱最终增 殖分化为合成和分泌各类免疫球蛋白分子的浆细泡。 B细胞可通过其抗原呈递作用刺激T细胞抗原受体(TCR/CD3)产生活化信号1,并通过其表面B7及其它协同刺激分子与T细陶表面的相应 受体分子如CD28.FA1等相结合,可激发产生协同刺激信号(即信号2),在双信号作用下可使TH细胞活化. B细胞要通过其BCR即表面g受体与抗原结合,g和g邮链与CD3分子相当,可传递活化信号1。近年证明在B细胞表面表达的CD40分子可 与T细胞表面的相应配体分子gP39(即CD40配体分子,CD40L)结合可激发其产生活化信号2,在此双信号作用下可使B细胞活化(图11, s)
CD8+T(50%) CDw49/CD29 (VLA-4、5、6) 异二聚体 粘合素 白细胞 其它细胞 细胞外基质VCAM-1 + + gP39(CD40L) CD4+T细胞 CD40 阻断或给予第2信号,可以人为调节免疫应答使之增强或或抑制,对免疫治疗提供了新的手段。如阻断第2信号的产生可使T细胞处于免疫 耐受状态,降低机体的免疫应答,这对防止移植排斥的发生和对超敏性疾病以及自身免疫性疾病的治疗是有利的。如将B7基因导入一定的肿瘤 细胞则可增强机体的抗肿瘤免疫应答,目前这方面的实验研究已有较多的报道(图11-4)。 图11-4 协同刺激信号与免疫应答调节 二、TH细胞与B细胞之间的相互作用 (一)B细胞对T细胞依赖抗原的应答 一般情况下当大量抗原进入未经免疫的机体后,诱发初次免疫应答时其抗原呈递细胞多由巨噬细胞完成。经Mφ活化TH细胞后再由活化的 TH细胞辅助B细胞产生抗体和形成记忆B细胞。但当再次免疫应答发生时,抗原呈递细胞则主要由已扩增的B细胞克隆承担。由于其膜Ig受体亲 和力增高故对少量抗原也能摄取,故可取代巨噬细胞的抗原呈递作用。 过去曾认为B细胞与TH细胞间的相互作用是通过抗原的桥联作用使之发生接触,即B细胞的抗原受体可识别抗原分子的半抗原部分,而TH 细胞的抗原受体可识别抗原分子的载体部分。但近年的实验证明B细胞与Mφ相同,也是通过抗原呈递作用与TH细胞相互作用。即B细胞可通过 其膜Ig受体蛋白质抗原,即T依赖性抗原结合,经内化作用将抗原摄入。然后经过加工处理,使T细胞决定簇与MHCⅡ类分子结合,形成MHC- 肽分子复合物并运送至B细胞表面,将其呈递给TH细胞,所以B细胞与TH细胞间的相互作用也是MHC限制性的。 B细胞和TH细胞通过其粘附分子及抗原呈递作用可彼此直接接触,并能相互诱导使之活化。活化B细胞在TH细胞的辅助下,B细胞最终增 殖分化为合成和分泌各类免疫球蛋白分子的浆细胞。 B细胞可通过其抗原呈递作用刺激T细胞抗原受体(TCR/CD3)产生活化信号1,并通过其表面B7及其它协同刺激分子与T细胞表面的相应 受体分子如CD28、IFA-1等相结合,可激发产生协同刺激信号(即信号2),在双信号作用下可使TH细胞活化。 B细胞要通过其BCR即表面Ig受体与抗原结合,Iga和Igβ链与CD3分子相当,可传递活化信号1。近年证明在B细胞表面表达的CD40分子可 与T细胞表面的相应配体分子gP39(即CD40配体分子,CD40L)结合可激发其产生活化信号2,在此双信号作用下可使B细胞活化(图11- 5)