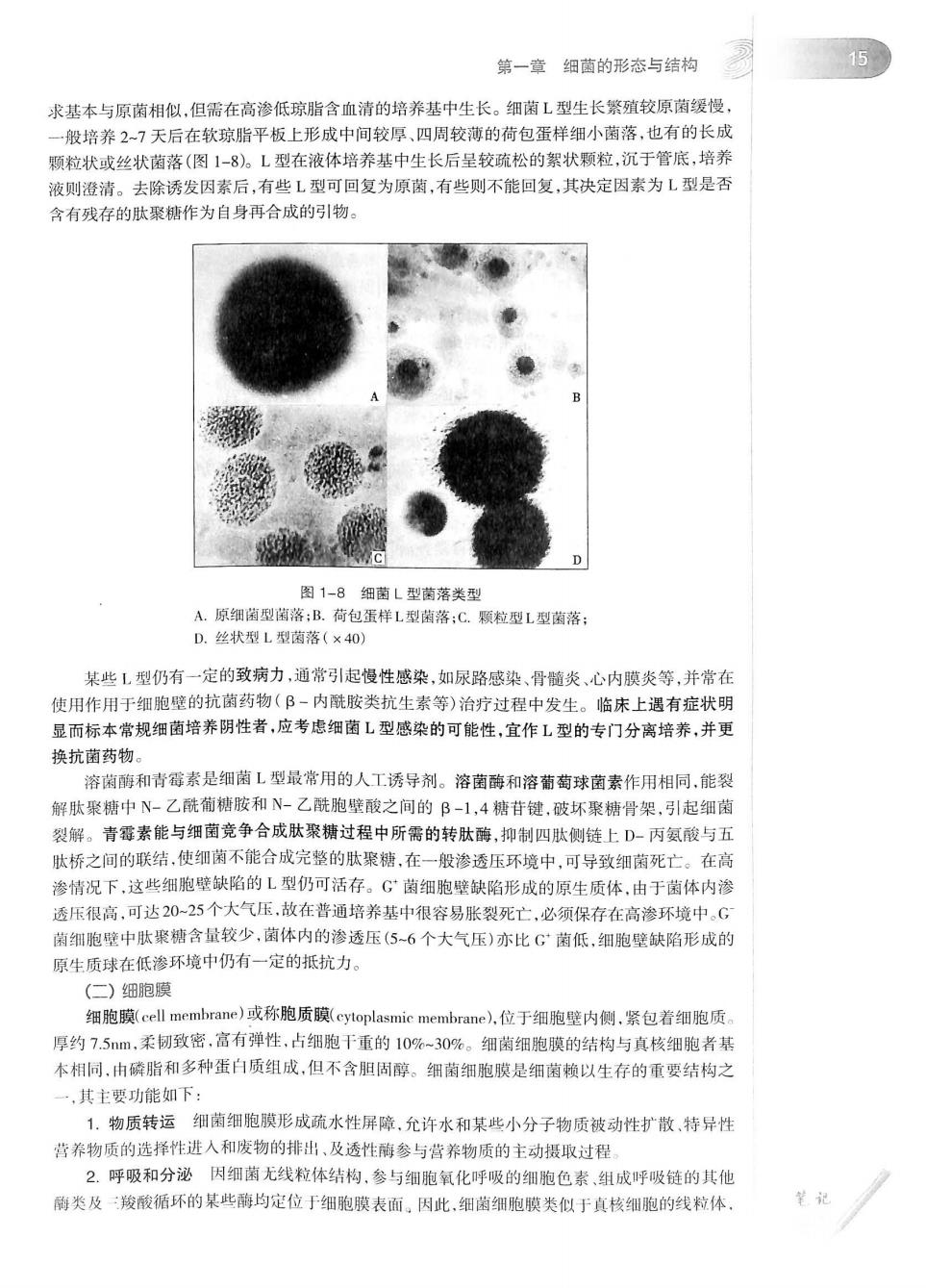
第一章细菌的形态与结构 15 求基本与原菌相似,但需在高渗低琼脂含血清的培养基中生长。细菌L型生长繁殖较原菌缓慢 一般培养27天后在软琼脂平板上形成中间较厚、四周较薄的荷包蛋样细小菌落,也有的长成 颗粒状或丝状菌落(图1-8。L型在液体培养基中生长后呈较硫松的絮状颗粒,沉于管底,培养 液则澄清。去除诱发因素后,有些L型可回复为原菌,有些则不能回复,其决定因素为L型是否 含有残存的肽聚糖作为自身再合成的引物 图1-8细菌L型黄落类型 A.原细菌型菌落:B.荷包蛋样L型菌落:C.颗粒型L型菌落 D.丝状型L型菌落(×40 某些1型仍有一定的致病力,通常引起慢性感染,如尿路感染、骨随炎、心内膜炎等,并常在 使用作用于细胞壁的抗菌药物(B-内酰胺类抗生素等)治疗过程中发生。临床上遇有症状明 显而标本常规细菌培养阴性者,应考虑细菌L型感染的可能性,宜作L型的专门分离培养,并更 换抗菌药物。 溶菌酶和青霉素是细菌L型最常用的人工诱导剂。溶菌酶和溶葡萄球菌素作用相同,能裂 解肽聚糖中N-乙酰葡糖胺和N-乙酰胞壁酸之间的B-1,4糖苷键,破环聚糖骨架,引起细菌 裂解。青霉素能与细菌竞争合成肽聚精过程中所需的转肽酶,抑制四肽侧链上D-丙氨酸与五 肽桥之间的联结,使细菌不能合成完整的肽聚糖,在一般渗透压环境中,可导致细菌死亡。在高 渗情况下,这些细胞壁缺陷的L型仍可活存。G菌细胞壁缺陷形成的原生质体,由于菌体内渗 透压很高,可达20-25个大气压,故在普通培养基中很容易胀裂死亡,必须保存在高渗环境中。( 菌细胞壁中肽聚糖含量较少,菌体内的渗透压(56个大气压)亦比G菌低,细胞壁缺陷形成的 原生质球在低渗环境中仍有一定的抵抗力。 (二)细胞膜 细胞膜(cell membrane)或称胞质膜(cytoplasmic membrane),位于细胞壁内侧,紧包若细胞质。 厚约7.5m,柔韧致密,高有弹性,占细胞干重的10%-309%。细菌细胞膜的结构与真核细胞者基 木相同,由磅脂和多种蛋白质组成,但不含胆周醇。细菌细胞膜是细菌赖以生存的重要结构之 ,其主要能如下: 1.物质转运细菌细胞膜形成硫水性屏障,允许水和某些小分子物质被动性扩散,特异性 营养物质的选择性进入和废物的排出、及透性酶参与营养物质的主动摄取过程 2,呼吸和分泌因细菌无线粒体结枸,参与细胞氧化呼吸的细胞色素组成呼吸链的其他 葡类及一羧酸循环的某些箭均定位于细胞膜表面。因此,细菌细胞膜类似于真核细胞的线粒体。 笔记

16 第一篇细苗学 在细胞呼吸和能量代谢中发挥重要作用。 3.生物合成细胞膜含有多种酶类,参 与细胞结构(如肽聚糖、磷脂、鞭毛和荚膜等 的合成。其中与肽聚糖合成有关的酶类(转肽 酶或转糖基酶),是青霉素作用的主要鞭位,称 为青霉素结合蛋白(penicillin-binding protein PBP),与细菌的耐药性形成有关 4.参与细菌分裂细菌部分细胞膜 内陷、折叠、卷曲形成的囊状物,称为中介体 (mesosome)。中介体多见于革兰阳性细菌(图 1-9),常位于菌体侧面(侧中介体)或靠近中部 (横膈中介体),可有一个或多个。中介体一端 图1-9白喉棒状杆菌的中介体 连在细胞膜上,另一端与核质相连,细胞分裂 (透射电镜×130000,谢念铭提供, 时中介体亦一分为二,各携一套核质进人子代细胞,有类似真核细胞纺锤丝的作用。中介体的形 成,有效地扩大了细胞膜面积,相应地增加了酶的含量和能量的产生,其功能类似于真核细胞的 线粒体,故亦称为拟线粒体(chondroid。 细菌的分泌系统是一种贯穿细菌胞膜的特殊结构,由不同的膜镶嵌蛋白构成。其分泌的底 物主要为蛋白质(如蛋白酶、溶血素、毒素等)和DA,可分布于细菌表面,或释放到细菌的外环 境中,或者注人宿主细胞内,参与细菌的各种重要生命活动和致病作用。如通过分泌系统,细菌 可将某些胞外酶分泌至胞外,消化营养物质,便于自身吸收利用:有些细菌蛋白可分泌到细胞膜 外,参与菌毛和鞭毛的生物合成:而分泌到细胞外的细菌毒素及毒性酶类,则参与细菌的致病过 程。根据细菌分泌系统的结构和功能不同,目前确认的有I~I型分泌系统。 ()细胞质 细胞膜包裹的溶胶状物质为细胞质(cytoplasm)或称原生质(protoplasm),由水,蛋白质,脂类、 核酸及少量糖和无机盐组成,其中含有许多重要结构 1.核糖体(b0s0m心)核糖体是细菌合成蛋白质的场所,游离存在于细胞质中,每个细菌体 内可达数万个。细菌核糖体沉降系数为70S,由50S和30S两个亚基组成,以大肠埃希菌为例 其化学组成66%是RNA(包括23S、16S和5SRNA),34%为蛋白质。核糖体常与正在转录的 mRNA相连呈“串珠”状,称多聚核糖体(polysome),使转录和转译偶联在一起。在生长活跃的细 菌体内,几乎所有的核糖体都以多聚核糖体的形式存在。 细菌的核糖体与真核生物核糖体不同,后者沉降系数为80S,由60S和40S两个亚基组成 有些抗生素如链鑫素能与细菌核精体的30S亚基结合,红春素与细菌核糖体的50S亚基结合 均能干扰其蛋白质合成,从而杀死细菌;但这些药物对人类的核糖体则无作 2.质粒(plasmid)质粒是细菌染色体外的遗传物质,存在于细胞质中。 为闭合环状的双链 DNA,带有遗传信息,控制细菌某些特定的遗传性状。质粒能独立自行复制,随细菌分裂转移到 子代细胞中。质粒不是细菌生长所必不可少的,失去质粒的细菌仍能正常存活。质粒除决定该 菌自身的某种性状外,还可通过接合或转导作用等将有关性状传递给另一细菌。质粒编码的细 菌性状有菌毛、细菌素、毒素和耐药性的产生等,与细菌致病性和耐药性有关。质粒的结构简单。 易导人细胞中,常作为载体广泛应用于生物学研究中。 3.胞质颗粒细菌细胞质中含有多种颗粒,大多为贮藏的营养物质,包括糖原,淀粉等多 糖脂类,磷酸盐等。胞质颗粒又称为内含物(inclusion),不是细菌的恒定结构,不同菌有不同的 胞质颗粒,同一菌在不同环境或生长期亦可不同。当营养充足时,胞质颗粒较多:养料和能源知 记 缺时,颗粒减少甚至消失。胞质颗粒中有一种主要成分是RNA和多偏磷酸盐(polymetaphosphate
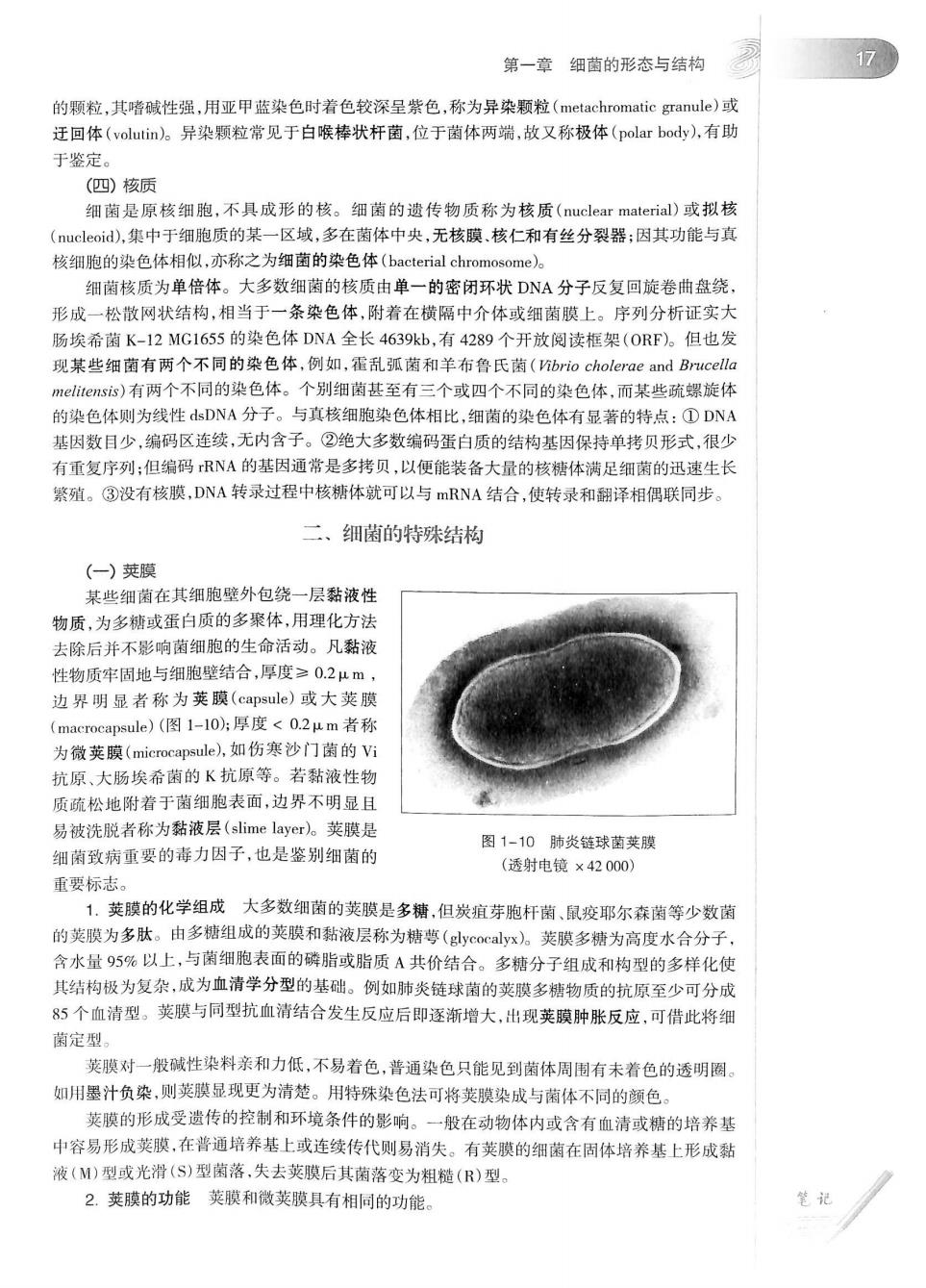
第一章细菌的形态与结构 17 的颗粒,其嗜碱性强,用亚甲蓝染色时着色较深呈紫色,称为异染颗粒(metachromatic granule)或 迁回体(volutin)。异染颗粒常见于白喉棒状杆菌,位于菌体两端,故又称极体(polar body),有助 于鉴定 (四核质 细菌是原核细胞,不具成形的核。细菌的遗传物质称为核质(nuclear material)或拟核 (),集中于细胞质的某一区域,多在菌体中央,无核膜,核仁和有丝分裂器:因其功能与真 核细跑的染色体相似,亦称之为细菌的染色体(bacterial chromosome)。 细菌核质为单倍体。大多数细菌的核质由单一的密闭环状DNA分子反复回旋卷曲:绕 形成一松散网状结构,相当于一条染色体,附着在横隔中介体或细菌膜上。序列分析证实大 肠埃希菌K-12MG1655的染色体DNA全长4639kb,有4289个开放阅读框架(ORF)。但也发 现某些细菌有两个不同的染色体,例如,霍乱弧菌和羊布鲁氏菌(brio cholerae and Brucelle melirensis))有两个不同的染色体。个别细菌甚至有三个或四个不同的染色体.而某些疏螺旋 的染色体则为线性dsDNA分子。与真核细胞染色体相比,细菌的染色体有显著的特点:①DNA 基因数目少,编码区连续,无内含子。②绝大多数编码蛋白质的结构基因保持单拷贝形式,很少 有重复序列:但编码RNA的基因通常是多拷贝,以便能装备大量的核糖体满足细菌的迅速生长 繁殖。③没有核膜,DNA转录过程中核糖体就可以与mRNA结合,使转录和翻译相偶联同步。 二、细菌的特殊结构 一)英膜 某些细菌在其细胞壁外包绕一层黏液性 物质,为多糖或蛋白质的多聚体,用理化方法 去除后并不影响菌细胞的生命活动。凡黏液 性物质牢固地与细胞壁结合,厚度≥0.2μm 边界明显者称为荚膜(capsule)或大荧膜 (macrocapsule)(图1-10片厚度<0.2μm者称 为微荚膜(miccapsule,)如伤寒沙门菌的V 抗原、大肠埃希菌的K抗原等。若黏液性物 质疏松地附若于菌细胞表面,边界不明显且 易被洗脱者称为黏液层(slime layer)。荚膜是 图1-10肺炎链球菌英膜 细南致病重要的毒力因子,也是鉴别细菌的 (透射电镜×42000) 重要标志。 1.荚膜的化学组成大多数细菌的荚膜是多糟,但炭疽芽胞杆菌、鼠疫耶尔森菌等少数菌 的荚膜为多肽。由多糖组成的荚膜和黏液层称为精尊(y0clx)》。荚膜多精为高度水合分子】 含水量95%以上,与菌细胞表面的磷脂或脂质A共价结合。多糖分子组成和构型的多样化使 其结构极为复杂,成为血清学分型的基础。例如肺炎链球菌的荚膜多糖物质的抗原至少可分成 85个血清型。荚膜与同型抗血清结合发生反应后即逐渐增大,出现荚膜肿胀反应,可借此将细 菊定型。 荚膜对一般碱性染料亲和力低,不易着色,普通染色只能见到菌体周围有未着色的透明圈。 如用墨汁负染,则荚膜显现更为清楚。用特殊染色法可将荚膜染成与菌体不同的颜色 英膜的形成受遗传的控制和环境条件的影响。一般在动物体内或含有血清或糖的培养基 中容易形成荚膜,在普通培养基上或连续传代则易消失。有荚膜的细菌在固体培养基上形成黏 液(M)型或光滑(S)型菌落,失去荚膜后其菌落变为粗糙(R)型。 2.荚膜的功能荚膜和微荚膜具有相同的功能。 笔记

18 第一篇细菌学 ()抗吞噬作用:荚膜具有保护细菌抵抗宿主吞噬细胞的吞噬和消化的作用,增强细菌的侵 袭力,因而荚膜是病原菌的重要毒力因子。荚膜多糖亲水和带负电荷,与吞噬细胞膜有静电排 斥力,赦能阻滞表面吞噬活性。例如肺炎链球菌,有荚膜株数个菌就可使实验小鼠致死,无荚膜 株则需上亿个菌才能使小鼠死亡。 (2)黏附作用:荚膜多糖可使细菌彼此粘连,也可黏附于组织细胞或无生命物体表面,参与 生物被膜(biofilm)的形成,是引起感染的重要因素。变异链球菌(S.mutans)依靠荚膜将其周定 在牙齿表面,利用口腔中的蔗糖产生大量的乳酸,积聚在附着部位形成生物被膜,导致牙齿珐琅 质的破坏,发生龋齿。有些具有荚膜细菌(例如铜绿假单胞菌),在住院病人的各种导管内黏附定 居形成生物被膜,是医院内感染发生的重要因素。 (③)抗有害物质的损伤作用:荚膜处于菌细胞的最外层,有保护菌体避免和减少受溶菌酶、 补体、抗体和抗菌药物等有害物质的损伤作用。 (一)手 许多细菌,包括所有的弧菌和螺菌,约半数的杆菌和个别球菌,在菌体上附有细长并呈波U 弯曲的丝状物,少仅1-2根,多者达数百根。这些丝状物称为鞭毛(1ag©lum),是细菌的运动器官 鞭毛长5-20μm,直径12-30mm,需用电子显微镜观察(图1-11),或经特殊染色法使鞭毛增粗后 才能在普通光学显微镜下看到(图1-12)。 图1-11破伤风梭菌的周身鞭毛 图1-12伤寒沙门菌的鞭毛 (透射电镜×16000,谢念铭提供) (鞭毛染色×1900) 根据鞭毛的数量和部位,可将鞭毛菌分成4类(图1-l3):①单毛菌(mono trichate),只有 一根鞭毛,位于菌休一端,如霍乱弧菌;②双毛菌(amphitrichate,)菌体两端各有一根鞭毛,如空 肠弯曲菌;③丛毛菌(ophoricha),菌体一端或两端有一丛鞭毛,如铜绿假单胞菌:④周毛菌 (peritrichate),菌体周身遍布许多鞭毛,如伤寒沙门菌。 1.鞭毛的结构鞭毛自细胞膜长出,游离于菌细胞外,由基础小体,钩状体和丝状体三个部 分组成(图1-14 鞭毛是从尖端生长,在菌体内形成的鞭毛蛋白分子不断地添加到鞭毛的末端。若用机械方 法去除鞭毛,新的鞭毛很快合成,3-6分钟内恢复动力。各菌种的鞭毛蛋白结构不同,具有高度 的抗原性,称为鞭毛(H)抗原。 2.狮手的功能 )为细菌的运动器官:具有鞭毛的细菌在液体环境中能主动、自由游动,速度迅速,如单细 毛的霍乱弧菌每秒移动可达55μm,周毛闲移动较慢,每秒25-30μm。细菌的运动有化学趋向 性,常向营养物质处前进,而逃离有害物质。 (2)有些细菌鞭毛与致病性有关:例如霍乱孤菌、空肠膏曲菌等通过活泼的鞭毛运动穿透小
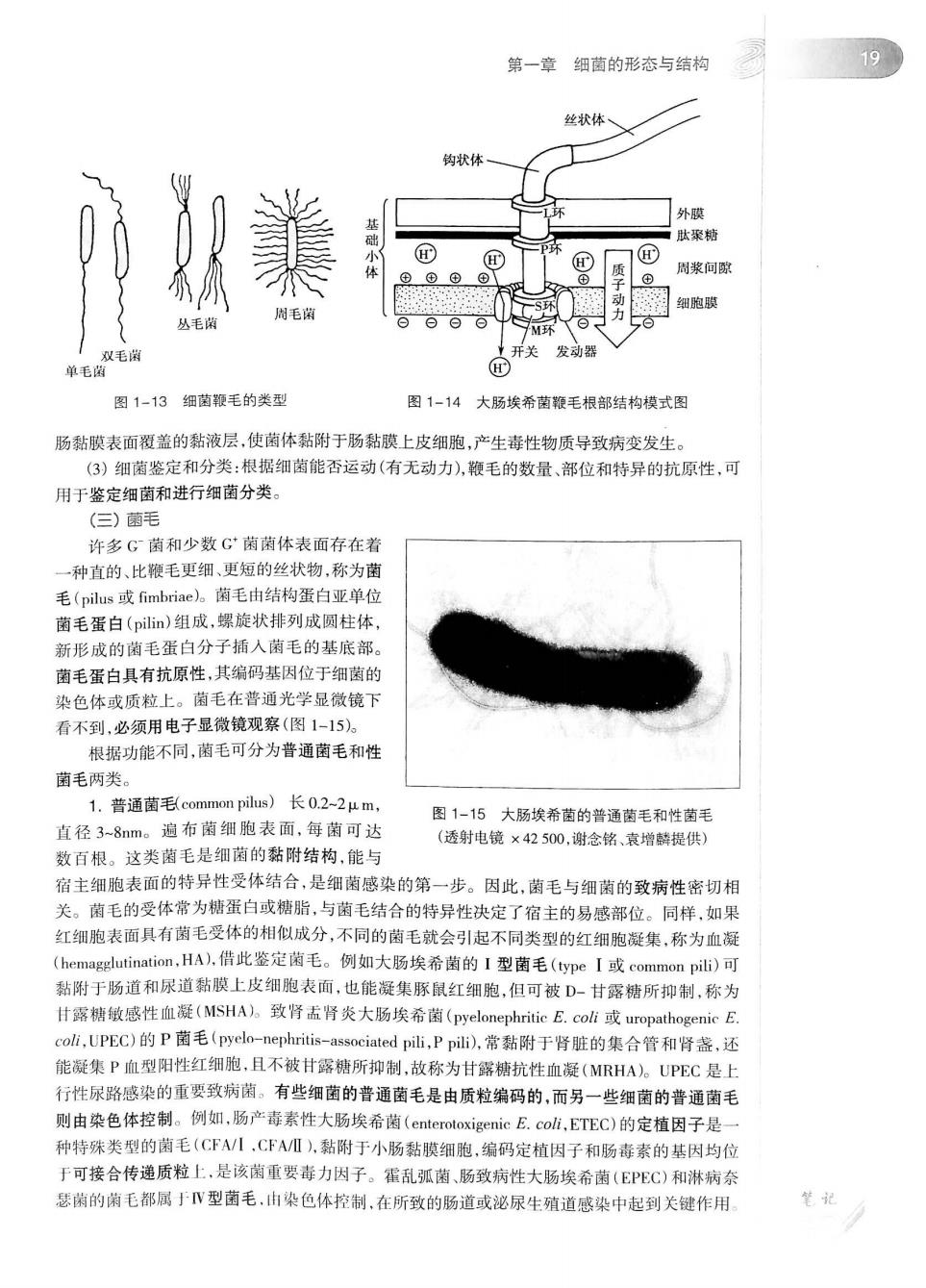
第一章细菌的形态与结构 19 丝状体、 钩状体 肽聚糖 周浆何 丛毛瑞 周毛闲 环 毛毛 开关发动器 图1-13细苗鞭毛的类型 图1-14大肠埃希菌鞭毛根部结构模式图 肠黏膜表面覆盖的黏液层,使菌体黏附于肠黏膜上皮细胞,产生毒性物质导致病变发生。 (3)细菌鉴定和分类:根据细菌能否运动(有无动力),鞭毛的数量、部位和特异的抗原性,可 用于鉴定细菌和进行细菌分类。 ()菌毛 许多G菌和少数G菌菌体表面存在着 一种直的、比瓶毛更细、更短的丝状物,称为黄 毛(pilus或fimbriae)。菌毛由结构蛋白亚单 菌毛蛋白(pilin)组成,螺旋状排列成圆柱体 新形成的菌毛蛋白分子插人菌毛的基底部】 菌毛蛋白具有抗原性,其编码基因位于细菌的 染色体或质粒上。菌毛在普通光学显微镜下 看不到,必须用电子显微镜观察(图1一15。 根据功能不同,菌毛可分为普通菌毛和性 菌毛两类。 1.普通菌毛)长02-2μm, 图1-15大肠埃希的普通菌毛和性菌毛 直径3-8nm。遍布菌细胞表面,每菌可达 (透射电镜×42500,谢念铭、袁增解提供) 数百根。这类菌毛是细菌的黏附结构,能与 宿主细胞表面的特异性受体结合,是细菌感染的第一步。因此,菌毛与细菌的致病性密切相 关。菌毛的受体常为糖蛋白或糖脂,与菌毛结合的特异性决定了宿主的易感部位。同样,如果 红细胞表面具有菌毛受体的相似成分,不同的菌毛就会引起不同类型的红细胞凝集,称为血凝 (hemagglutination,HA),件此鉴定菌毛。例如大肠埃希菌的I型菌毛(type I或common pili)可 黏附于肠道和尿道黏膜上皮细胞表面,也能凝集豚鼠红细胞,但可被D-甘露精所抑制,称为 甘露糖敏感性血凝(MSHA)。致肾去肾炎大肠埃希菌(pyelonephritieE,coi或uropathogenic E coli,UPEC)的P菌毛(pye-ephritis-associated pili,P pili)),常黏附于肾脏的集合管和肾盏,还 能凝集P血型阳性红细胞,且不被甘露糖所抑制,放称为甘露糖抗性血凝(MRHA)。UPEC是上 行性尿路感染的重要致病菌。有些细菌的普通菌毛是由质粒编码的,而另一些细菌的普通菌毛 则由染色体控制。例如,肠产毒素性大肠埃希菌(enteroloxigenie.coi,ETEC)的定植因子是 种特殊类型的菌毛(CFV1,CFAI),黏附于小肠黏膜细胞,编码定植因子和肠毒素的基因均位 于可接合传递质粒上,是该菌重要毒力因子。霍乱弧菌、肠致病性大肠埃希菌(EPEC)和淋病命 瑟菊的菌毛都属于V型菌毛,由染色体控制,在所致的肠道或泌尿生殖道感染中起到关键作用