
真核生物细胞内存在两种甲基化酶活性:一 种被称为日常型(mainte-nance)甲基转移酶, 另一种是 从头合成(denovo synthesis)甲基转 移酶。前者主要在甲基化母链(模板链)指导下 使处于半甲基化的DNA双链分子上与甲基胞嘧啶 相对应的胞嘧啶甲基化。日常型甲基转移酶常常 与DNA内切酶活性相耦联,有3种类型。II类酶活 性包括内切酶和甲基化酶两种成分,而I类和III类 都是双功能酶,既能将半甲基化DNA甲基化,又 能降解外源无甲基化DNA
真核生物细胞内存在两种甲基化酶活性:一 种被称为日常型(mainte-nance)甲基转移酶, 另一种是 从头合成(denovo synthesis)甲基转 移酶。前者主要在甲基化母链(模板链)指导下 使处于半甲基化的DNA双链分子上与甲基胞嘧啶 相对应的胞嘧啶甲基化。日常型甲基转移酶常常 与DNA内切酶活性相耦联,有3种类型。II类酶活 性包括内切酶和甲基化酶两种成分,而I类和III类 都是双功能酶,既能将半甲基化DNA甲基化,又 能降解外源无甲基化DNA

CHa ACGTATCGT ACGTATE methylated cytosine me助ation mthated 1106 iiW 5 cytosine CHa TGCATAGCA TGCATAE ACGTATCGT 3 oreced by 3 6 DNA y mainterance maintenan图 TGCATAGCA tepleation methe 简h链 CHa ACGTATCGT ACGTATC 3 ii田 methylation 5 ii册 5 TGCATAGCA TGCATAR 4-68 C

表7-2细胞内3类日常型甲基转移酶活性分析 特性 1类 1类 1类 蛋白质结构 甲基化与内切酶功能 双功能醇,2个亚基 双功能酶,3个亚基 由不同蛋白质执行 识别位点 4~6bp,常为回文结构 5~1bp,常为不对称序列由两个不对称序列构成 酶切位点 同识别位点或非常靠 识别位点下游24~25bp 非特异性,可在距识别 近该位点 位点>1000bp处 内切/甲基化相互独立 同时进行 不能同时进行 其他 不需要ATP水解 需要ATP水解 需要ATP水解

由于甲基化胞嘧啶极易在进化中丢失,所以,高等 真核生物中CG序列远远低于其理论值。哺乳类基因组 中约存在4万个CG islands,大多位于转录单元的5‘区。 没有甲基化的胞嘧啶发生脱氨基作用,就可能被氧 化成为U,被DNA修复系统所识别和切除,恢复成C。 已经甲基化的胞嘧啶发生脱氨基作用, 它就变为T, 无法 被区分。因此, CpG序列极易丢失
由于甲基化胞嘧啶极易在进化中丢失,所以,高等 真核生物中CG序列远远低于其理论值。哺乳类基因组 中约存在4万个CG islands,大多位于转录单元的5‘区。 没有甲基化的胞嘧啶发生脱氨基作用,就可能被氧 化成为U,被DNA修复系统所识别和切除,恢复成C。 已经甲基化的胞嘧啶发生脱氨基作用, 它就变为T, 无法 被区分。因此, CpG序列极易丢失
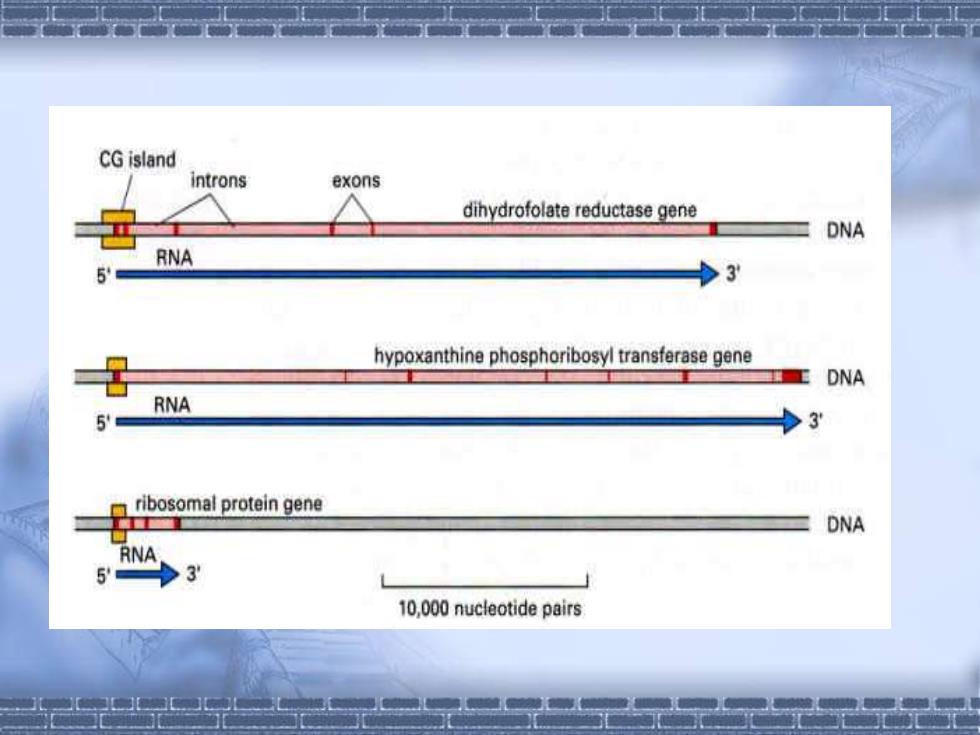
CG island introns exons dihydrofolate reductase gene DNA RNA 5= >3 hypoxanthine phosphoribosyl transferase gene DNA RNA 5■ >3 ribosomal protein gene DNA RNA 5 10,000 nucleotide pairs