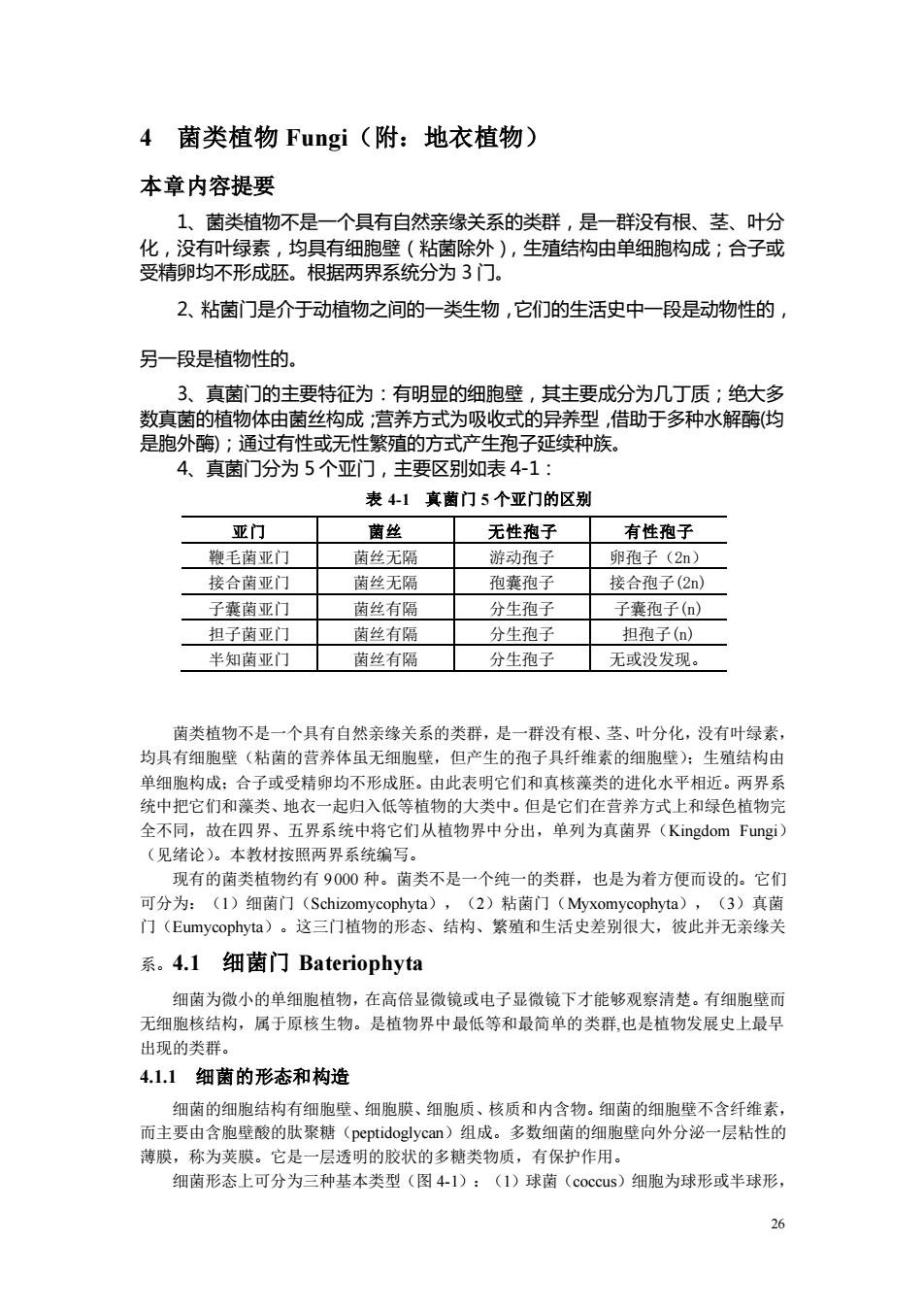
4菌类植物Fungi(附:地衣植物) 本章内容提要 1、菌类植物不是一个具有自然亲缘关系的类群,是一群没有根、茎、叶分 化,没有叶绿素,均具有细胞壁(粘菌除外),生殖结构由单细胞构成;合子或 受精卵均不形成胚。根据两界系统分为3门。 2、粘菌门是介于动植物之间的一类生物,它们的生活史中一段是动物性的, 另一段是植物性的, 3、真菌门的主要特征为:有明显的细胞壁,其主要成分为几丁质;绝大多 数真菌的植物体由菌丝构成:营养方式为吸收式的异养型,借助于多种水解酶(均 是胞外酶);通过有性或无性繁殖的方式产生孢子延续种族。 4、真菌门分为5个亚门,主要区别如表4-1: 表41真菌门5个亚门的区别 亚门 菌丝 无性孢子 有性孢子 鞭毛菌亚门 菌丝无隔 游动孢子 卵孢子(2n) 接合南亚门 菌丝无 孢囊孢子 接合孢子(2n) 子囊南亚门 菌丝有隔 分生孢子 子囊孢子(n) 担子菌亚门 菌丝有隔 分生孢子 担孢子(n) 半知南亚门☐菌丝有隔 分生孢子 无或没发现。 菌类植物不是一个具有自然亲缘关系的类群,是一群没有根、茎、叶分化,没有叶绿素 均具有细胞壁(粘菌的营养体虽无细胞壁,但产生的孢子具纤维素的细胞壁):生殖结构由 单细胞构成:合子或受精卵均不形成胚,由此表明它们和真核藻类的进化水平相近。两界系 统中把它们和藻类、地衣一起归入低等植物的大类中。但是它们在营养方式上和绿色植物完 全不同,故在四界、五界系统中将它们从植物界中分出,单列为真菌界(Kingdom Fungi) (见绪论)。本教材按照两界系统编写。 现有的菌类植物约有9000种。菌类不是一个纯一的类群,也是为着方便而设的。它们 可分为:(1)细茵门(Schiz ,(2)粘菌门(M (3)真茵 系。4.1细菌门Bateriophyta 细菌为微小的单细胞植物,在高倍显微镜或电子显微镜下才能够观察清楚。有细胞壁而 无细胞核结构,属于原核生物。是植物界中最低等和最简单的类群也是植物发展史上最昂 出现的类群。 4.1.1细菌的形态和构造 细菌的细胞结构有细胞壁、细胞膜、细胞质、核质和内含物。细菌的细胞壁不含纤维素, 而主要由含胞壁酸的肽聚糖(peptidoglycan)组成。多数细菌的细胞壁向外分泌一层粘性的 薄膜,称为荚膜。它是一层透明的胶状的多糖类物质,有保护作用。 细南形态上可分为三种基本类型(图41):(I)球菌(coccus)细胞为球形或半球形
26 4 菌类植物 Fungi(附:地衣植物) 本章内容提要 1、菌类植物不是一个具有自然亲缘关系的类群,是一群没有根、茎、叶分 化,没有叶绿素,均具有细胞壁(粘菌除外),生殖结构由单细胞构成;合子或 受精卵均不形成胚。根据两界系统分为 3 门。 2、粘菌门是介于动植物之间的一类生物,它们的生活史中一段是动物性的, 另一段是植物性的。 3、真菌门的主要特征为:有明显的细胞壁,其主要成分为几丁质;绝大多 数真菌的植物体由菌丝构成;营养方式为吸收式的异养型,借助于多种水解酶(均 是胞外酶);通过有性或无性繁殖的方式产生孢子延续种族。 4、真菌门分为 5 个亚门,主要区别如表 4-1: 表 4-1 真菌门 5 个亚门的区别 亚门 菌丝 无性孢子 有性孢子 鞭毛菌亚门 菌丝无隔 游动孢子 卵孢子(2n) 接合菌亚门 菌丝无隔 孢囊孢子 接合孢子(2n) 子囊菌亚门 菌丝有隔 分生孢子 子囊孢子(n) 担子菌亚门 菌丝有隔 分生孢子 担孢子(n) 半知菌亚门 菌丝有隔 分生孢子 无或没发现。 菌类植物不是一个具有自然亲缘关系的类群,是一群没有根、茎、叶分化,没有叶绿素, 均具有细胞壁(粘菌的营养体虽无细胞壁,但产生的孢子具纤维素的细胞壁);生殖结构由 单细胞构成;合子或受精卵均不形成胚。由此表明它们和真核藻类的进化水平相近。两界系 统中把它们和藻类、地衣一起归入低等植物的大类中。但是它们在营养方式上和绿色植物完 全不同,故在四界、五界系统中将它们从植物界中分出,单列为真菌界(Kingdom Fungi) (见绪论)。本教材按照两界系统编写。 现有的菌类植物约有 9 000 种。菌类不是一个纯一的类群,也是为着方便而设的。它们 可分为:(1)细菌门(Schizomycophyta),(2)粘菌门(Myxomycophyta),(3)真菌 门(Eumycophyta)。这三门植物的形态、结构、繁殖和生活史差别很大,彼此并无亲缘关 系。4.1 细菌门 Bateriophyta 细菌为微小的单细胞植物,在高倍显微镜或电子显微镜下才能够观察清楚。有细胞壁而 无细胞核结构,属于原核生物。是植物界中最低等和最简单的类群,也是植物发展史上最早 出现的类群。 4.1.1 细菌的形态和构造 细菌的细胞结构有细胞壁、细胞膜、细胞质、核质和内含物。细菌的细胞壁不含纤维素, 而主要由含胞壁酸的肽聚糖(peptidoglycan)组成。多数细菌的细胞壁向外分泌一层粘性的 薄膜,称为荚膜。它是一层透明的胶状的多糖类物质,有保护作用。 细菌形态上可分为三种基本类型(图 4-1):(1)球菌(coccus)细胞为球形或半球形

直径0.5-2μm,没有鞭毛,不能运动。(2)杆菌(bacillus)细胞呈杆棒状,长1.5-10μm, 究0.5-1μm,通常具鞭毛,能游动。(3)螺旋菌(spirillum)细胞长而弯曲,略略弯曲的 称为弧菌,通 具鞭 能游动 细菌的营养方式多数为异养,有的是从活的动植物体内吸收有机物,称寄生细菌。有的 是从动植物遗体或其它有机物取得有机物,称腐生细菌。但也有少数细菌是自养的,如光合 细菌(photosynthetic bacteria)和化能自养细菌(chemoautotrophic bacteria)。光合细菌体内 含细黄叶绿素,其分子结不同与蓝藻和直核细附的叶绿素,吸收被长范用在660一870 的近红外区,能够利 光合作 用直 利用光能合成有机物:而化能自养细菌则是通过氧化无 机物(如NH、HhS等)而获得能量 4.1.2细菌的繁殖 细菌的繁殖方式以细胞分裂方式进行,无有性生殖。繁殖时,细胞壁的中部 向内凹入,在凹入处生长出新细胞壁,把细胞分成两个,所以细菌也常常被称为 "裂殖菌”(Schizomycetes 细菌的裂殖速度极快,在适宜的条件下,20~30分钟就能分裂一次,并可 继续分裂若干次。细菌分裂时,需要充分的营养和一定的温度等,因此,细菌的 分裂常常受外界条件的限制。在不利的条件下,某些细菌生失水浓缩,形成1个 圆形或椭圆形的内生孢子,称为芽孢(endospore,芽孢的壁厚,渗透性很弱, 含水少,能抵抗不良的环境,芽孢可存活十几年,当遇到适宜的环境,可再产生 新的菌体。由于一个细胞仅形成一个芽孢,所以它并无繁殖功能,而是渡过不良 环境的一种适应结构。 细菌分布广,几乎分布在地球的各个角落。其中有些细菌能导致严重的疾病,如霍乱 破伤风、猩红热、伤寒、鼠疫、结核等。尽管如此,细菌在自然界的生态系统中具有不可替 代的作用,它是物质循环中不可缺少的一员,同时细菌在农业(如生物固氨)和工业(如细 菌发酵)生产中也具有十分重要的作用。 4.2粘菌门Myxomycophyta 4.2.1粘菌门的一般特征 粘菌门是介于动植物之间的一类生物,约有500种。它们的生活史中一段是动物性的, 另一段是植物性的 粘菌在生长期或营养期为裸的无细胞噬、多核的原生质团,称变形体(: odium) 其营养体的构造、运动和摄食的方式,与原生动物的变形虫很相似,具有运动性的特点。但 在繁殖时营固着生活,能产生具有纤维素细胞壁的孢子,又具有植物性的特点
27 直径 0.5-2μm,没有鞭毛,不能运动。(2)杆菌(bacillus)细胞呈杆棒状,长 1.5-10μm, 宽 0.5-1μm,通常具鞭毛,能游动。(3)螺旋菌(spirillum)细胞长而弯曲,略略弯曲的 称为弧菌,通常具鞭毛,能游动。 细菌的营养方式多数为异养,有的是从活的动植物体内吸收有机物,称寄生细菌。有的 是从动植物遗体或其它有机物取得有机物,称腐生细菌。但也有少数细菌是自养的,如光合 细菌(photosynthetic bacteria)和化能自养细菌(chemoautotrophic bacteria)。光合细菌体内 含细菌叶绿素,其分子结构不同与蓝藻和真核细胞的叶绿素,吸收波长范围在 660-870nm 的近红外区,能够利用光合作用直接利用光能合成有机物;而化能自养细菌则是通过氧化无 机物(如 NH3、H2S 等)而获得能量。 4.1.2 细菌的繁殖 细菌的繁殖方式以细胞分裂方式进行,无有性生殖。繁殖时,细胞壁的中部 向内凹入,在凹入处生长出新细胞壁,把细胞分成两个,所以细菌也常常被称为 “裂殖菌”(Schizomycetes)。 细菌的裂殖速度极快,在适宜的条件下,20~30分钟就能分裂一次,并可 继续分裂若干次。细菌分裂时,需要充分的营养和一定的温度等,因此,细菌的 分裂常常受外界条件的限制。在不利的条件下,某些细菌生失水浓缩,形成1个 圆形或椭圆形的内生孢子,称为芽孢(endospore)。芽孢的壁厚,渗透性很弱, 含水少,能抵抗不良的环境,芽孢可存活十几年,当遇到适宜的环境,可再产生 新的菌体。由于一个细胞仅形成一个芽孢,所以它并无繁殖功能,而是渡过不良 环境的一种适应结构。 细菌分布广,几乎分布在地球的各个角落。其中有些细菌能导致严重的疾病,如霍乱、 破伤风、猩红热、伤寒、鼠疫、结核等。尽管如此,细菌在自然界的生态系统中具有不可替 代的作用,它是物质循环中不可缺少的一员,同时细菌在农业(如生物固氮)和工业(如细 菌发酵)生产中也具有十分重要的作用。 4.2 粘菌门 Myxomycophyta 4.2.1 粘菌门的一般特征 粘菌门是介于动植物之间的一类生物,约有 500 种。它们的生活史中一段是动物性的, 另一段是植物性的。 粘菌在生长期或营养期为裸露的无细胞壁、多核的原生质团,称变形体(plasmodium), 其营养体的构造、运动和摄食的方式,与原生动物的变形虫很相似,具有运动性的特点。但 在繁殖时营固着生活,能产生具有纤维素细胞壁的孢子,又具有植物性的特点

4.2.1粘菌门的代表植物 发网菌属(Stemont)是粘菌中最常见的种类,其变形体呈不规则网状,直径数厘米, 能借助体形的改变在阴湿处的腐木上或枯叶上缓慢爬行,并能吞食固体食物。在繁殖时,变 形虫爬到干燥光亮的地方,形成很多的发状突起(图42),每个突起发育成一个具柄的孢子 囊(子实体):孢子囊通常为长筒形,外有包被(peridium),孢子囊柄伸入囊内的部分称囊轴 (columella),囊内有孢丝(capillitium)交织成网:原生质团中的许多核同时进行减数分裂, 进而原生质团制裂成许多块单核的小原生质 每块小原生质先分泌出细胞壁,形成一孢了 藏在孢丝的网眼中,成熟时,包被破裂,借助于包网的弹力把孢子弹出:孢子在适合的环境 中即可萌发为具两条不等长鞭毛的游动细胞,游动细胞的鞭毛可以收缩,使游动细胞变成 个变形体状细胞,称变形菌胞(myxamoebae):由游动细胞或变形菌胞两两配合,形成合子, 合子核不经过休眠即进行多次有丝分裂,形成多数二倍体核,构成一个多核的变形体。 4.3真菌门Eumycophyta 4.3.1真菌的一般特征 4.3.1.1营养体 直菌(下ng)属其核异养生物,点黄的细胞内不含叶绿素,也没有质体,营寄生或腐生生 活。真菌贮存的养分主要是肝糖(liver starch),还有少量的蛋白质、脂肪以及微量的维生素 真菌多数种类有明显的细胞壁,其主要成分为几丁质(chitin)和纤维素(cellulose)),一般低等 真菌的细胞壁多由纤维素组成,而高等真菌以几丁质为主。 除少数单细胞真菌(如酵母)外,绝大多数真菌的植物体由菌丝(hyphae)构成,菌丝是纤 细的管状体,有无隔菌丝和有隔菌丝之分。无隔菌丝是一个长管形细胞,有分枝或无,大多 数是多核的,低等真菌 的菌丝 般为无隔菌丝,仅在受伤或产生生殖结构时才 产生全封闭的 隔膜(图43):有隔菌丝中有隔膜把菌丝隔成许多细胞,每个细胞内含1或2个核,高等 真菌的菌丝多为有隔菌丝。但菌丝中的横隔上通常有各种类型小孔,原生质甚至核可以经小 孔流通。横隔上的小孔主要有3种类型(图43):单孔型、多孔型和桶孔式,桶孔式隔膜 的结构最为复杂,隔膜中央有1孔,但孔的边缘增厚膨大成桶状,并在两边的孔外各有1个 由内质网形成的弧形膜,称桶孔覆垫或隔膜孔帽, 真菌主要利用菌丝吸收养分,腐生菌可由菌丝直接从基质中吸收养分,也可产生假根 (hiz0id)用于吸收养分:寄主细胞内寄生的真菌通过直接与寄生细胞的原生质接触而吸收养 分。胞间寄生的真菌则利用从菌丝体上特化产生的吸器(haustorium)(图4-4)伸入寄主细胞 内吸取养料。吸取养料的过程是首先借助于多种水解酶(均是胞外酶),把大分子物质分解为 可溶性的小分子物质,然后借助于较高的渗透压吸收。寄生真菌的渗透压一般比寄主高2 5倍 腐生菌的渗 更高 真菌在繁殖或环境条件不良时,菌丝常相互密结, 形成两种组织:拟薄壁组 (pseudoparenchyma)和疏丝组织(prosenchyma),再构成菌丝组织体,常变态为三种形态 (图4-5):根状茵索(rhizomorph):菌丝体密结呈绳索状,外形似根。子座(stroma):容 纳子实体的辉座,是从营养阶段到繁殖阶段的过渡形式。菊核(clerotium):由菌丝密结成 须色、质地坚硬的核状体。子实体 )也是一种菌丝组织体,为含有或产生孢 子的组织结构。能形子实体的真,人们称为大型真。 4.3.1.2真菌的繁殖 真菌繁殖的方式多种多样,并涉及很多不同类型的孢子(图46)。少数单细胞真菌如
28 4.2.1 粘菌门的代表植物 发网菌属(Stemonitis)是粘菌中最常见的种类,其变形体呈不规则网状,直径数厘米, 能借助体形的改变在阴湿处的腐木上或枯叶上缓慢爬行,并能吞食固体食物。在繁殖时,变 形虫爬到干燥光亮的地方,形成很多的发状突起(图 4-2),每个突起发育成一个具柄的孢子 囊(子实体);孢子囊通常为长筒形,外有包被(peridium),孢子囊柄伸入囊内的部分称囊轴 (columella),囊内有孢丝(capillitium)交织成网;原生质团中的许多核同时进行减数分裂, 进而原生质团割裂成许多块单核的小原生质,每块小原生质先分泌出细胞壁,形成一孢子, 藏在孢丝的网眼中,成熟时,包被破裂,借助于包网的弹力把孢子弹出;孢子在适合的环境 中即可萌发为具两条不等长鞭毛的游动细胞,游动细胞的鞭毛可以收缩,使游动细胞变成一 个变形体状细胞,称变形菌胞(myxamoebae);由游动细胞或变形菌胞两两配合,形成合子, 合子核不经过休眠即进行多次有丝分裂,形成多数二倍体核,构成一个多核的变形体。 4.3 真菌门 Eumycophyta 4.3.1 真菌的一般特征 4.3.1.1 营养体 真菌(Fungi)属真核异养生物,真菌的细胞内不含叶绿素,也没有质体,营寄生或腐生生 活。真菌贮存的养分主要是肝糖(liver starch),还有少量的蛋白质、脂肪以及微量的维生素。 真菌多数种类有明显的细胞壁,其主要成分为几丁质(chitin)和纤维素(cellulose),一般低等 真菌的细胞壁多由纤维素组成,而高等真菌以几丁质为主。 除少数单细胞真菌(如酵母)外,绝大多数真菌的植物体由菌丝(hyphae)构成,菌丝是纤 细的管状体,有无隔菌丝和有隔菌丝之分。无隔菌丝是一个长管形细胞,有分枝或无,大多 数是多核的,低等真菌的菌丝一般为无隔菌丝,仅在受伤或产生生殖结构时才产生全封闭的 隔膜(图4-3);有隔菌丝中有隔膜把菌丝隔成许多细胞,每个细胞内含 1 或 2 个核,高等 真菌的菌丝多为有隔菌丝。但菌丝中的横隔上通常有各种类型小孔,原生质甚至核可以经小 孔流通。横隔上的小孔主要有 3 种类型(图4-3):单孔型、多孔型和桶孔式,桶孔式隔膜 的结构最为复杂,隔膜中央有 1 孔,但孔的边缘增厚膨大成桶状,并在两边的孔外各有 1 个 由内质网形成的弧形膜,称桶孔覆垫或隔膜孔帽。 真菌主要利用菌丝吸收养分,腐生菌可由菌丝直接从基质中吸收养分,也可产生假根 (rhizoid)用于吸收养分;寄主细胞内寄生的真菌通过直接与寄生细胞的原生质接触而吸收养 分。胞间寄生的真菌则利用从菌丝体上特化产生的吸器(haustorium)(图4-4)伸入寄主细胞 内吸取养料。吸取养料的过程是首先借助于多种水解酶(均是胞外酶),把大分子物质分解为 可溶性的小分子物质,然后借助于较高的渗透压吸收。寄生真菌的渗透压一般比寄主高 2~ 5 倍,腐生菌的渗透压更高。 真菌在繁殖或环境条件不良时,菌丝常相互密结,形成两种组织:拟薄壁组织 (pseudoparenchyma)和疏丝组织(prosenchyma),再构成菌丝组织体,常变态为三种形态 (图 4-5):根状菌索(rhizomorph):菌丝体密结呈绳索状,外形似根。子座(stroma):容 纳子实体的褥座,是从营养阶段到繁殖阶段的过渡形式。菌核(sclerotium):由菌丝密结成 颜色深、质地坚硬的核状体。子实体(sporophore)也是一种菌丝组织体,为含有或产生孢 子的组织结构。能形成子实体的真菌,人们称为大型真菌。 4.3.1.2 真菌的繁殖 真菌繁殖的方式多种多样,并涉及很多不同类型的孢子(图4-6)。少数单细胞真菌如

裂殖酵母南属(S$ohi00 ©s)主要通过细胞分裂立生子细胞,而大部分直南可以通 过产生芽生孢子、厚孢子或节孢子等进行营养紧殖。芽生孢子《 e)是从一个细 胞出芽形成 挂生 脱离母 后即长成 个新个体 厚壁孢 y1 菌丝中个别细胞膨大形成的休眠孢子,其原生质浓缩细胞壁加厚,渡过不良环境后,再萌 发为菌丝体:节孢子(arthrospore)是由菌丝细胞断裂形成的。 真菌无性生殖也极为发达,并在无性生殖过程中也形成多种不同类型的孢子,包括游动 孢子、孢囊孢子和分生孢子等。游动孢子(z00s Ore)是水生真菌产生的借水传播的孢子,无 壁,具鞭毛,能游动,在游动孢子囊( rangiu 孢囊孢 在孢子囊(sporangium)内形成的不动孢子,借气流传播:分生孢子(conidiospore)是由分生孢 子囊梗的项端或侧面产生的一种不动孢子,借气流或动物传播。 真菌的有性生殖方式也极其多样化,有些真菌可产生单细胞的配子,以同配或异配的方 式进行有性生殖:另有一些真菌通过两性配子囊的结合形成“合子”,这种类型的合子习 惯上称之为接合孢子 )或的子( 子套茵有性配合后, 形成子 ,在子 结合后产生的孢子,和无性生殖的孢子完全不同。 真菌通过各种途径产生的孢子在适宜的环境条件下萌发,生长形成菌丝体(myc©lium), 精坐体在一个生长季里可产生若干代无性孢子,这是生活史的无性阶段:真菌在生长的后 期,常形成配子囊,产生配子 一般先行质配,形成双核细胞,再行核配,形成合子:通 常合子形成后很快即进行减数分裂,形成单倍的孢子,再萌发成单倍的菌丝体 这样就完 成了 个生活周期。由此可见,在真南的生活史中,二倍体时期只是很短暂的合子阶段, 合子是一个细胞而不是一个营养体,所以,大多数真菌的生活史中,只有核相交替,而没 有世代交替。 4.3.2分类及代表植物 真菌是生物界中很大的一个类群,约12万种,通常分为5个亚门,即鞭毛菌亚门纲, 接合菌亚门、子囊菌亚门、担子菌亚门和半知菌亚门。 4.3.2.1鞭毛菌亚门(Mastigomycotina) 少数低等种类营养体为单细胞,大多数为无隔多核、分技的丝状体,细胞壁的成分为纤 维素或几丁质。繁殖时繁殖器官的基部产生隔膜,形成孢子囊或配子囊。无性生殖产生具有 鞭毛的游动孢子:有性生殖时产生卵孢子或休眠孢子 水霉属(Saprolegnia),寄生在鱼类或其他水生动物的尸体上,孢子通过鱼的伤口萌发 并将菌丝穿入组织中,吸收寄主的养料,使其致死。菌丝体呈白色,无性繁殖时,菌体顶端 影大,多数细胞核向这里流动,然后膨大基部产生横隔,形成一个长筒形的游动孢子囊,霸 中产生孢子。孢子从顶端小孔处游出,然后在旧孢子囊基部再生出第二个,如此重复称层出 形成(现象)。游动孢子梨形,顶生2条鞭毛,称初生孢子,后变为肾形,侧生2条鞭毛的 游动孢子,称次生孢子,这种现象为双游现象(diplanetism) 有性繁殖时,在菌丝顶端形成精囊和卵囊,精子和卵细胞结合形成2倍体的卵孢子,卵 孢子经过休眠,萌发时首先进行减数分裂,然后形成新的菌丝体。水霉属的生活史见图47: 4.3.2.2接合菌亚门(亿ygomycotina) 营养体为为无隔多核、分枝的菌丝组成的菌丝体,细胞壁的成分为几丁质和壳聚糖。有 的种类产生厚垣孢子和节孢子进行营养繁殖:无性生殖产生孢囊孢子:有性生殖产生接合孢 子。 29
29 裂殖酵母菌属(Sohizosaccaromyces)主要通过细胞分裂产生子细胞,而大部分真菌可以通 过产生芽生孢子、厚壁孢子或节孢子等进行营养繁殖。芽生孢子(blastospore)是从一个细 胞出芽形成的,芽生孢子脱离母体后即长成一个新个体;厚壁孢子(chlamydospore)是由 菌丝中个别细胞膨大形成的休眠孢子,其原生质浓缩,细胞壁加厚,渡过不良环境后,再萌 发为菌丝体;节孢子(arthrospore)是由菌丝细胞断裂形成的。 真菌无性生殖也极为发达,并在无性生殖过程中也形成多种不同类型的孢子,包括游动 孢子、孢囊孢子和分生孢子等。游动孢子(zoospore)是水生真菌产生的借水传播的孢子,无 壁,具鞭毛,能游动,在游动孢子囊(zoosporangium)中形成;孢囊孢子(sporangiospore)是 在孢子囊(sporangium)内形成的不动孢子,借气流传播;分生孢子(conidiospore)是由分生孢 子囊梗的顶端或侧面产生的一种不动孢子,借气流或动物传播。 真菌的有性生殖方式也极其多样化,有些真菌可产生单细胞的配子,以同配或异配的方 式进行有性生殖;另有一些真菌通过两性配子囊的结合形成“合子”,这种类型的合子习 惯上称之为接合孢子(zygospore)或卵孢子(oospore)。子囊菌有性配合后,形成子囊,在子 囊内产生子囊孢子。担子菌有性生殖后,在担子上形成担孢子。担孢子和子囊孢子是有性 结合后产生的孢子,和无性生殖的孢子完全不同。 真菌通过各种途径产生的孢子在适宜的环境条件下萌发,生长形成菌丝体(mycelium), 菌丝体在一个生长季里可产生若干代无性孢子,这是生活史的无性阶段;真菌在生长的后 期,常形成配子囊,产生配子,一般先行质配,形成双核细胞,再行核配,形成合子;通 常合子形成后很快即进行减数分裂,形成单倍的孢子,再萌发成单倍的菌丝体,这样就完 成了一个生活周期。由此可见,在真菌的生活史中,二倍体时期只是很短暂的合子阶段, 合子是一个细胞而不是一个营养体,所以,大多数真菌的生活史中,只有核相交替,而没 有世代交替。 4.3.2 分类及代表植物 真菌是生物界中很大的一个类群,约 12 万种,通常分为 5 个亚门,即鞭毛菌亚门纲、 接合菌亚门、子囊菌亚门、担子菌亚门和半知菌亚门。 4.3.2.1 鞭毛菌亚门(Mastigomycotina) 少数低等种类营养体为单细胞,大多数为无隔多核、分枝的丝状体,细胞壁的成分为纤 维素或几丁质。繁殖时繁殖器官的基部产生隔膜,形成孢子囊或配子囊。无性生殖产生具有 鞭毛的游动孢子;有性生殖时产生卵孢子或休眠孢子。 水霉属(Saprolegnia),寄生在鱼类或其他水生动物的尸体上,孢子通过鱼的伤口萌发 并将菌丝穿入组织中,吸收寄主的养料,使其致死。菌丝体呈白色,无性繁殖时,菌体顶端 膨大,多数细胞核向这里流动,然后膨大基部产生横隔,形成一个长筒形的游动孢子囊,囊 中产生孢子。孢子从顶端小孔处游出,然后在旧孢子囊基部再生出第二个,如此重复称层出 形成(现象)。游动孢子梨形,顶生 2 条鞭毛,称初生孢子,后变为肾形,侧生 2 条鞭毛的 游动孢子,称次生孢子,这种现象为双游现象(diplanetism)。 有性繁殖时,在菌丝顶端形成精囊和卵囊,精子和卵细胞结合形成 2 倍体的卵孢子,卵 孢子经过休眠,萌发时首先进行减数分裂,然后形成新的菌丝体。水霉属的生活史见图 4-7。 4.3.2.2 接合菌亚门(Zygomycotina) 营养体为为无隔多核、分枝的菌丝组成的菌丝体,细胞壁的成分为几丁质和壳聚糖。有 的种类产生厚垣孢子和节孢子进行营养繁殖;无性生殖产生孢囊孢子;有性生殖产生接合孢 子

黑根需属(Rhi仁Ops)亦称面包霉,是一种常见的腐生菌,常生长在馒头、面包、水果 其菌丝体由无隔菌丝组成, ,分枝多: 黑根霉的菌丝在基质表 与假根相对处,向上产生若干直立的菌丝,随 发育的进行,直立菌丝的项端膨大形成孢子囊,孢子囊的中央有一个半球形的囊轴 (codumela),囊轴和囊壁之间产生许多孢囊孢子,孢囊孢子成熟时呈黑色,孢子囊破裂后, 响子散出并萌发成新的南丝体(图48)。 黑根霉有性生殖为异宗配合,两个不同宗的菌丝在外形上很难分辨,因此常用“+” “-”号加以区分:进行有性生 殖时 ”菌丝各产生 一短枝,短枝顶端膨大形成 配子囊:当“+ ”配子囊相互接触时,接触处的壁脸解消失, 两个配 子囊的原生质混合,细胞核成对地融合,形成一个具多个二倍体核的接合孢子。接合孢了 成熟时黑色,具疣状突起,休眠后,在适宜的条件下萌发产生一个孢子囊梗,其顶端形成 接合孢子囊,其内的二倍体核经减数分裂产生多个单倍的“+” “”孢子:接合孢子 囊壁破裂 孢子散出 ,分别萌发成新的菌丝体 根霉属用途很广,它含有大量的淀粉酶能分解淀粉为葡萄糖,这种糖化作用是酿酒的 第一步,酿酒的第二步是酵母菌的发酵作用,把葡萄糖发酵为酒。 4.3.2.3子囊菌亚门(Ascomycotina) 子囊黄亚门是真黄门中种类最名的亚门。本亚门除酵母菌为单细跑外,绝大部分为有隔 茵丝组成的菌丝体:细胞壁的成分为几丁质。繁殖时,单细胞种类以出芽进行繁殖,多细 种类产生分生孢子:有性生殖时形成子囊,合子在子囊内进行减数分裂,形成子囊孢子 单细胞种类,子囊裸露,不形成子实体,多细胞种类形成子实体,子囊包于子实体内。 子实体是产生和容纳有性孢子的组织结构,子囊菌的子实体又称为子囊果(ascocarp, scoma),子囊果是由子实层和包被两部分构成。子囊果有3种类型:(1)闭囊壳 《cleistothecium),子里是球形.无引口,完全闭合 (2)子囊壳(perithecium) 囊果呈瓶形,顶端有孔口,这种子囊果常埋于子座中。 ( 子盘(apothecium 果呈盘状、杯状、碗状,子实层常露在外(图49)。 子囊果的形状为子囊菌亚门的重要分 类依据。火丝菌属(Pyronema)的菌丝为有隔菌丝,多分枝,菌丝体白色,棉絮状。无性生 殖是以分枝菌丝顶端产生分生孢子来完成。 有性生殖时,一些菌丝的顶端细胞膨大,形成具多核的精子囊(spom atangium)和的囊 特 产囊休 (ascogonium) 产囊体顶端有一条弯管状的受精丝(trichogyne】 其基部有 横隔与产囊体隔开:当精子囊与受精丝接触时,接触处细胞壁融成 小孔,此时受精丝基 的横隔也同时融化,于是,精子囊中的细胞质和雄核通过受精丝流入产囊体中,进行质配 雌雄核成对地排列:此后,经过有性过程的刺激,产囊体的上半部产生许多管状的产囊丝 (ascogenous-hypha),雌、雄核成对地移入产囊蚀中:随后,产囊独分枝并产生描壁形成多 个细胞,每一细胞中具一对核 囊丝顶端双核细胞伸长,并弯曲形成钩状体(产囊丝 钩,crosier) 双核同时分裂,形成4个核,此时钩状体产生横隔,隔成3个细胞:钩状 尖端细胞称钩尖,具一核,居中的细胞称钩头,具一对核,即子囊母细胞(ascus mother cell), 钩状体基部的细胞为钩柄,也具一核。子囊母细胞中的雌、雄核经核配后形成二倍的合子, 合子随即进行一次成数分裂和一次有丝分裂形成8个子核,之后经跑质分裂形成8个细胞 8个子孢子。潮若子查孢子的形成,子查母细胞也深渐变成状的且8个线形排列子 囊孢子的子囊(ascu 钩尖细胞经 钩细胞接 又形成 核细胞,此细胞再经过钩状构成(hook permation)重新形成新的子囊,如此反复进行形成多 数子囊。在子囊形成的同时,不有的产囊丝和一些营养菌丝在子囊之间形成侧丝(隔丝, paraphysis),子囊果内的子囊和侧丝排列成子实层(hymenium),另有大量的营养菌丝立 刻从立壶体的下方生出,除一部分参与形成测丝外,大部分形成子壶果的句被,句被和子实
30 黑根霉属(Rhizopus)亦称面包霉,是一种常见的腐生菌,常生长在馒头、面包、水果、 蔬菜或腐败的食物上。其菌丝体由无隔菌丝组成,多核,分枝多;黑根霉的菌丝在基质表 面匍匍生长,以假根伸入基质吸收养料;与假根相对处,向上产生若干直立的菌丝,随着 发育的进行,直立菌丝的顶端膨大形成孢子囊,孢子囊的中央有一个半球形的囊轴 (codumela),囊轴和囊壁之间产生许多孢囊孢子,孢囊孢子成熟时呈黑色,孢子囊破裂后, 孢子散出并萌发成新的菌丝体(图4-8)。 黑根霉有性生殖为异宗配合,两个不同宗的菌丝在外形上很难分辨,因此常用“+”、 “-”号加以区分;进行有性生殖时,“+”、“-”菌丝各产生一短枝,短枝顶端膨大形成 配子囊;当“+”、“-”配子囊相互接触时,接触处的壁融解消失,“+”、“-”两个配 子囊的原生质混合,细胞核成对地融合,形成一个具多个二倍体核的接合孢子。接合孢子 成熟时黑色,具疣状突起,休眠后,在适宜的条件下萌发产生一个孢子囊梗,其顶端形成 一接合孢子囊,其内的二倍体核经减数分裂产生多个单倍的“+”、“-”孢子;接合孢子 囊壁破裂后,孢子散出,分别萌发成新的菌丝体。 根霉属用途很广,它含有大量的淀粉酶能分解淀粉为葡萄糖,这种糖化作用是酿酒的 第一步,酿酒的第二步是酵母菌的发酵作用,把葡萄糖发酵为酒。 4.3.2.3 子囊菌亚门(Ascomycotina) 子囊菌亚门是真菌门中种类最多的亚门。本亚门除酵母菌为单细胞外,绝大部分为有隔 菌丝组成的菌丝体;细胞壁的成分为几丁质。繁殖时,单细胞种类以出芽进行繁殖,多细胞 种类产生分生孢子;有性生殖时形成子囊,合子在子囊内进行减数分裂,形成子囊孢子。 单细胞种类,子囊裸露,不形成子实体,多细胞种类形成子实体,子囊包于子实体内。 子实体是产生和容纳有性孢子的组织结构,子囊菌的子实体又称为子囊果(ascocarp, ascoma),子囊果是由子实层和包被两部分构成。子囊果有 3 种类型:(1)闭囊壳 (cleistothecium),子囊果呈球形,无孔口,完全闭合。(2)子囊壳(perithecium),子 囊果呈瓶形,顶端有孔口,这种子囊果常埋于子座中。(3)子囊盘(apothecium),子囊 果呈盘状、杯状、碗状,子实层常露在外(图 4-9)。子囊果的形状为子囊菌亚门的重要分 类依据。火丝菌属(Pyronema)的菌丝为有隔菌丝,多分枝,菌丝体白色,棉絮状。无性生 殖是以分枝菌丝顶端产生分生孢子来完成。 有性生殖时,一些菌丝的顶端细胞膨大,形成具多核的精子囊(spormatangium)和卵囊, 特称为产囊体(ascogonium),产囊体顶端有一条弯管状的受精丝(trichogyne),其基部有一 横隔与产囊体隔开;当精子囊与受精丝接触时,接触处细胞壁融成一小孔,此时受精丝基部 的横隔也同时融化,于是,精子囊中的细胞质和雄核通过受精丝流入产囊体中,进行质配, 雌雄核成对地排列;此后,经过有性过程的剌激,产囊体的上半部产生许多管状的产囊丝 (ascogenous-hypha),雌、雄核成对地移入产囊丝中;随后,产囊丝分枝并产生横壁形成多 个细胞,每一细胞中具一对核;继而产囊丝顶端双核细胞伸长,并弯曲形成钩状体(产囊丝 钩,crosier),双核同时分裂,形成 4 个核,此时钩状体产生横隔,隔成 3 个细胞;钩状体 尖端细胞称钩尖,具一核,居中的细胞称钩头,具一对核,即子囊母细胞(ascus mother cell), 钩状体基部的细胞为钩柄,也具一核。子囊母细胞中的雌、雄核经核配后形成二倍的合子, 合子随即进行一次减数分裂和一次有丝分裂形成 8 个子核,之后经胞质分裂形成 8 个细胞, 即 8 个子囊孢子。随着子囊孢子的形成,子囊母细胞也逐渐变成棒状的具 8 个线形排列子 囊孢子的子囊(ascus);与此同时,钩尖细胞经下弯与钩柄细胞接触、融合,又形成一个双 核细胞,此细胞再经过钩状构成(hook permation)重新形成新的子囊,如此反复进行形成多 数子囊。在子囊形成的同时,不育的产囊丝和一些营养菌丝在子囊之间形成侧丝(隔丝, paraphysis),子囊果内的子囊和侧丝排列成子实层(hymenium),另有大量的营养菌丝立 刻从产囊体的下方生出,除一部分参与形成侧丝外,大部分形成子囊果的包被,包被和子实