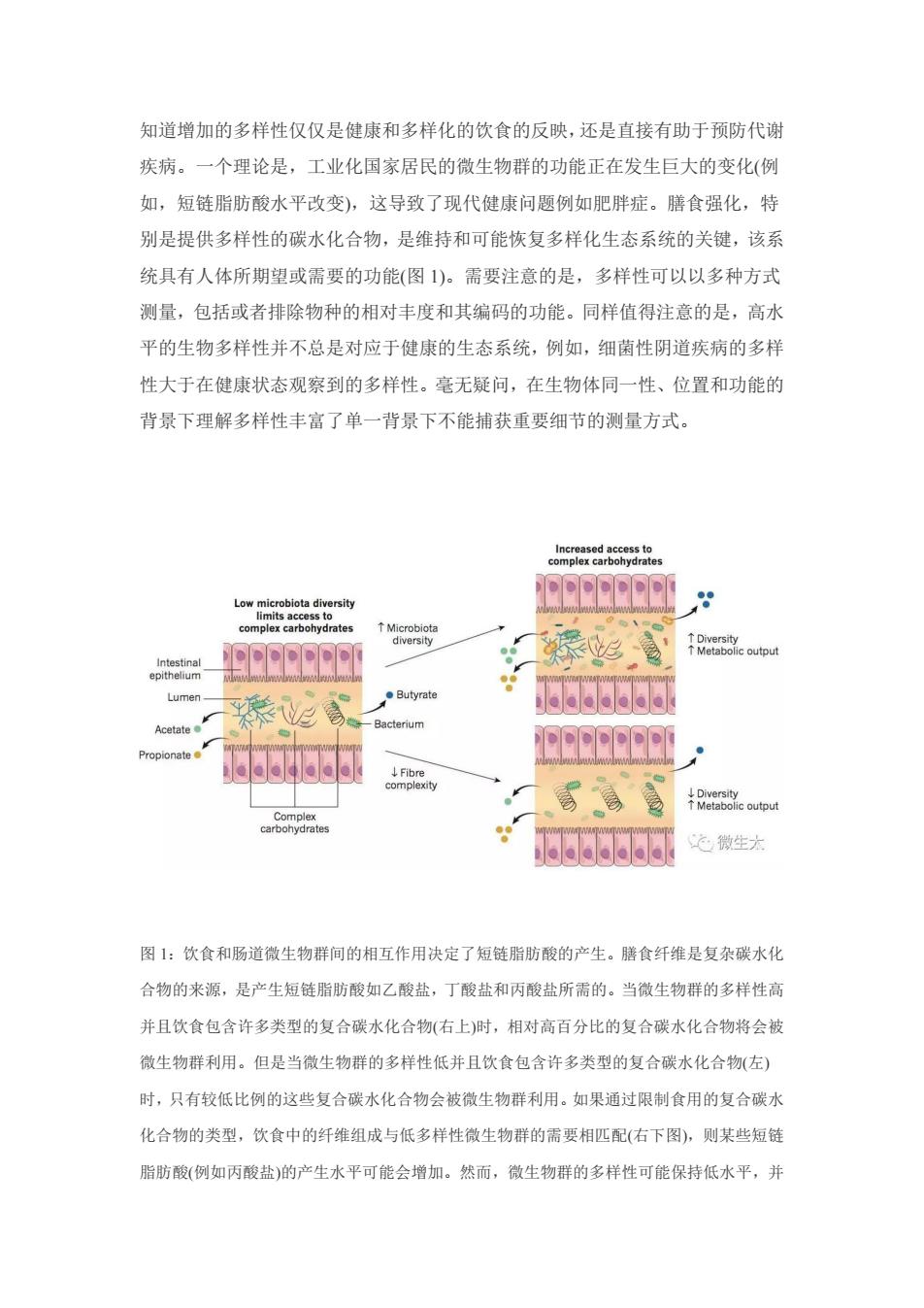
知道增加的多样性仅仅是健康和多样化的饮食的反映,还是直接有助于预防代谢 疾病。一个理论是,工业化国家居民的微生物群的功能正在发生巨大的变化(例 如,短链脂肪酸水平改变),这导致了现代健康问题例如肥胖症。膳食强化,特 别是提供多样性的碳水化合物,是维持和可能恢复多样化生态系统的关键,该系 统具有人体所期望或需要的功能(图1)。需要注意的是,多样性可以以多种方式 测量,包括或者排除物种的相对丰度和其编码的功能。同样值得注意的是,高水 平的生物多样性并不总是对应于健康的生态系统,例如,细菌性阴道疾病的多样 性大于在健康状态观察到的多样性。毫无疑问,在生物体同一性、位置和功能的 背景下理解多样性丰富了单一背景下不能捕获重要细节的测量方式。 congeedb6ne 99 drat EEREE ouput Lumer 高 66666d Propionate e taboc output 运微生大 图1:饮食和肠道微生物群间的相互作用决定了短链脂肪酸的产生。腾食纤维是复杂碳水化 合物的来源,是产生短链脂肪酸如乙酸盐,丁酸盐和丙酸盐所需的。当微生物群的多样性高 并且饮食包含许多类型的复合碳水化合物(右上)时,相对高百分比的复合碳水化合物将会被 微生物群利用。但是当微生物群的多样性低并且饮食包含许多类型的复合碳水化合物(左) 时,只有较低比例的这些复合碳水化合物会被微生物群利用。如果通过限制食用的复合碳水 化合物的类型,饮食中的纤维组成与低多样性微生物群的需要相匹配(右下图),则某些短链 脂肪酸(例如丙酸盐)的产生水平可能会增加。然而,微生物群的多样性可能保持低水平,并
知道增加的多样性仅仅是健康和多样化的饮食的反映,还是直接有助于预防代谢 疾病。一个理论是,工业化国家居民的微生物群的功能正在发生巨大的变化(例 如,短链脂肪酸水平改变),这导致了现代健康问题例如肥胖症。膳食强化,特 别是提供多样性的碳水化合物,是维持和可能恢复多样化生态系统的关键,该系 统具有人体所期望或需要的功能(图 1)。需要注意的是,多样性可以以多种方式 测量,包括或者排除物种的相对丰度和其编码的功能。同样值得注意的是,高水 平的生物多样性并不总是对应于健康的生态系统,例如,细菌性阴道疾病的多样 性大于在健康状态观察到的多样性。毫无疑问,在生物体同一性、位置和功能的 背景下理解多样性丰富了单一背景下不能捕获重要细节的测量方式。 图 1:饮食和肠道微生物群间的相互作用决定了短链脂肪酸的产生。膳食纤维是复杂碳水化 合物的来源,是产生短链脂肪酸如乙酸盐,丁酸盐和丙酸盐所需的。当微生物群的多样性高 并且饮食包含许多类型的复合碳水化合物(右上)时,相对高百分比的复合碳水化合物将会被 微生物群利用。但是当微生物群的多样性低并且饮食包含许多类型的复合碳水化合物(左) 时,只有较低比例的这些复合碳水化合物会被微生物群利用。如果通过限制食用的复合碳水 化合物的类型,饮食中的纤维组成与低多样性微生物群的需要相匹配(右下图),则某些短链 脂肪酸(例如丙酸盐)的产生水平可能会增加。然而,微生物群的多样性可能保持低水平,并

且可能不能提供类似于多样化微生物群的功能。那么食用复合饮食可能引起多种类型的短链 脂肪酸水平的增加(右上),有助于额外促进肠道微生物群的多样性。丙酸盐产生的水平与肠 道中拟杆菌种类的丰度相关,这与拟杆菌类参与丙酸盐产生的结论相一致。研究表明结肠中 的纤维发酵使pH降低,这可以帮助增加肠道微生物群的多样性或使某些类群增强了有利于 其自身生长的pH环境。 4微生物生态系统的底物 在膳食纤维中发现的许多植物多糖的结构是复杂的。因此,修饰,释放,运 输和代谢成分性单糖所需的许多酶不由人类基因组编码也属于正常。此外,这些 反应可能并不是发生在这些物质快速通过小肠时(简单的碳水化合物就是在此被 消化吸收的)。因此,复杂的碳水化合物被转移到达远端肠道,通过其密集的微 生物群体进行发酵。 根据实验室试验研究发现,许多复杂的植物碳水化合物可以作为膳食纤维。 然而,可以代谢的纤维的量(例如,通过糖苷键的酶促降解和释放的单糖发酵成 短链脂肪酸)将取决于许多因素,包括微生物群的组成。可被微生物群代谢的碳 水化合物被称为微生物群可利用的碳水化合物,可以与经过了消化道但不被代谢 的碳水化合物形成对照。这种代谢利用率是一个重要的差异特征,它将碳水化合 物定义为促进肠道种间生态经济的资源,意味着会产生代谢产物,如短链脂肪酸。 值得注意的是,微生物群的高度多样性与Burkina Faso地区农民中高水平的 短链脂肪酸产生量相关,也与符猎者中复杂碳水化合物代谢相关的微生物组基因 的富集相关。在小鼠的多代研究中,与富含微生物群可利用的碳水化合物的饮食 相比,西方饮食加剧了微生物群多样性的丧失,这与预期的糖苷水解酶多样性降 低相对应。几项人体研究表明,微生物群多样性和代谢产量存在特异性“上限”。 例如,在至少6个月的纯素食饮食或10天的高纤维低脂肪饮食之后,不足以使 微生物群多样性显著增加或产生粪便短链脂肪酸。植物性饮食可以显著改变肠道 微生物群的组成,尽管没有观察到多样性的变化。当抗性淀粉摄入量高时, Ruminococcus bromi及其同种类菌丰度未见增加,且在粪便中未消化的淀粉含量
且可能不能提供类似于多样化微生物群的功能。那么食用复合饮食可能引起多种类型的短链 脂肪酸水平的增加(右上),有助于额外促进肠道微生物群的多样性。丙酸盐产生的水平与肠 道中拟杆菌种类的丰度相关,这与拟杆菌类参与丙酸盐产生的结论相一致。研究表明结肠中 的纤维发酵使 pH 降低,这可以帮助增加肠道微生物群的多样性或使某些类群增强了有利于 其自身生长的 pH 环境。 4 微生物生态系统的底物 在膳食纤维中发现的许多植物多糖的结构是复杂的。因此,修饰,释放,运 输和代谢成分性单糖所需的许多酶不由人类基因组编码也属于正常。此外,这些 反应可能并不是发生在这些物质快速通过小肠时(简单的碳水化合物就是在此被 消化吸收的)。因此,复杂的碳水化合物被转移到达远端肠道,通过其密集的微 生物群体进行发酵。 根据实验室试验研究发现,许多复杂的植物碳水化合物可以作为膳食纤维。 然而,可以代谢的纤维的量(例如,通过糖苷键的酶促降解和释放的单糖发酵成 短链脂肪酸)将取决于许多因素,包括微生物群的组成。可被微生物群代谢的碳 水化合物被称为微生物群可利用的碳水化合物,可以与经过了消化道但不被代谢 的碳水化合物形成对照。这种代谢利用率是一个重要的差异特征,它将碳水化合 物定义为促进肠道种间生态经济的资源,意味着会产生代谢产物,如短链脂肪酸。 值得注意的是,微生物群的高度多样性与 Burkina Faso 地区农民中高水平的 短链脂肪酸产生量相关,也与狩猎者中复杂碳水化合物代谢相关的微生物组基因 的富集相关。在小鼠的多代研究中,与富含微生物群可利用的碳水化合物的饮食 相比,西方饮食加剧了微生物群多样性的丧失,这与预期的糖苷水解酶多样性降 低相对应。几项人体研究表明,微生物群多样性和代谢产量存在特异性“上限”。 例如,在至少 6 个月的纯素食饮食或 10 天的高纤维低脂肪饮食之后,不足以使 微生物群多样性显著增加或产生粪便短链脂肪酸。植物性饮食可以显著改变肠道 微生物群的组成,尽管没有观察到多样性的变化。当抗性淀粉摄入量高时, Ruminococcus bromii 及其同种类菌丰度未见增加,且在粪便中未消化的淀粉含量