
生理上克服不亲和性的可能途径: 1)花粉蒙导 (mentor pollen)法 在授不亲和花粉的同时,混入一些杀死的亲和花粉,蒙骗柱头, 从而达到受粉的目的。 2)物理化学处理法 采用变温、辐射、激素或抑制剂处理雌蕊组织,以打破不亲和性。 3)重复授粉 超量授粉,干扰识别反应 (1)破坏识别物质或抑制识别反应
生理上克服不亲和性的可能途径: 1)花粉蒙导 (mentor pollen)法 在授不亲和花粉的同时,混入一些杀死的亲和花粉,蒙骗柱头, 从而达到受粉的目的。 2)物理化学处理法 采用变温、辐射、激素或抑制剂处理雌蕊组织,以打破不亲和性。 3)重复授粉 超量授粉,干扰识别反应 (1)破坏识别物质或抑制识别反应

1)组织培养 利用胚珠、子房培养,试管受精。 2)细胞杂交 原生质体融合或转基因技术 1)蕾期授粉法 雌蕊识别蛋白尚未形成,雄性生殖单位可能成熟 2)延期授粉法 柱头内不亲和物质减少,活性减弱,对花粉萌发, 花粉管生长抑制作用降低 (2)避开雌蕊中识别物质的活性期 (3)去除识别反应组织
1)组织培养 利用胚珠、子房培养,试管受精。 2)细胞杂交 原生质体融合或转基因技术 1)蕾期授粉法 雌蕊识别蛋白尚未形成,雄性生殖单位可能成熟 2)延期授粉法 柱头内不亲和物质减少,活性减弱,对花粉萌发, 花粉管生长抑制作用降低 (2)避开雌蕊中识别物质的活性期 (3)去除识别反应组织

3 花粉萌发与花粉管生长 花粉萌发与花粉管生长表现出集体效应(group effect), 即落在柱头上花粉密度越大,萌发比例越高,花粉管生长越快。 原因:花粉中存在生长素,花粉数量越多,生长素也就越多, 所以促进花粉的萌发和花粉管的生长。 花粉为什么能向着胚囊定向生长? 由花粉管的向化性运动引起的。 雌蕊组织中产生“向化性物质”控制花粉管的可塑性; 同时,雌蕊组织中向化性物质分布的浓度不同,花粉管尖端朝 着向化性物质浓度递增的方向(柱头→胚囊)定向延伸-Ca2+
3 花粉萌发与花粉管生长 花粉萌发与花粉管生长表现出集体效应(group effect), 即落在柱头上花粉密度越大,萌发比例越高,花粉管生长越快。 原因:花粉中存在生长素,花粉数量越多,生长素也就越多, 所以促进花粉的萌发和花粉管的生长。 花粉为什么能向着胚囊定向生长? 由花粉管的向化性运动引起的。 雌蕊组织中产生“向化性物质”控制花粉管的可塑性; 同时,雌蕊组织中向化性物质分布的浓度不同,花粉管尖端朝 着向化性物质浓度递增的方向(柱头→胚囊)定向延伸-Ca2+
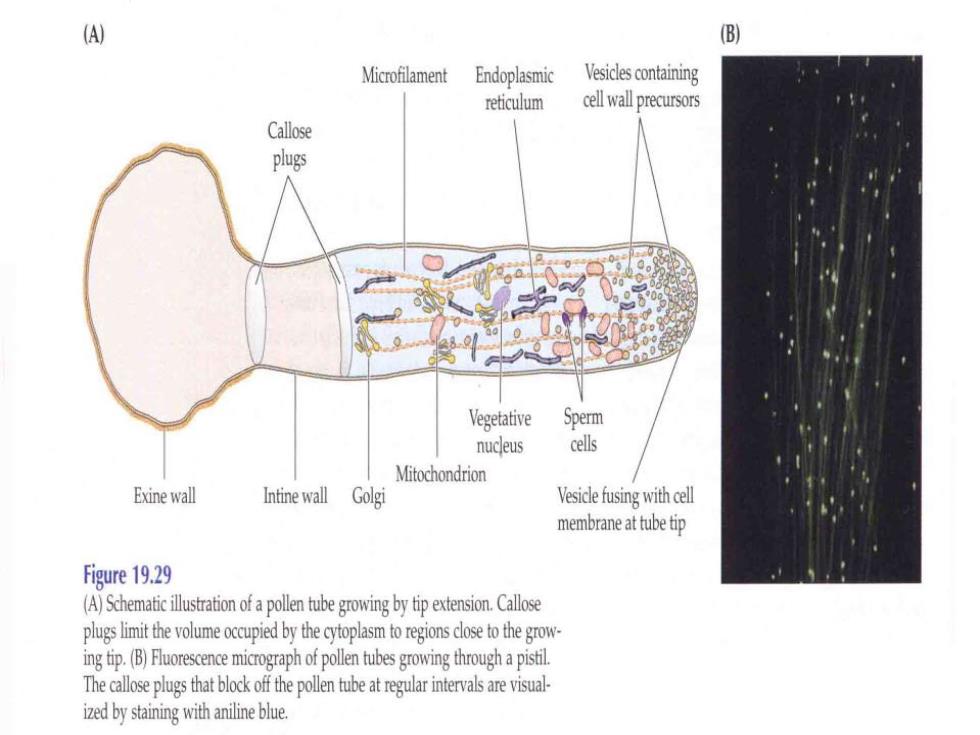
B Microfilament Endoplasmic Vesicles containing reticulum cell wall precursors Callose plugs Vegetative Sperm nucleus cells Mitochondrion Exine wall Intine wall Golgi Vesicle fusing with cell membrane at tube tip Figure 19.29 (A)Schematic illustration of a pollen tube growing by tip extension.Callose plugs limit the volume occupied by the cytoplasm to regions close to the grow ing tip.(B)Fluorescence micograph of pollen tubes growing through a pistil The callose plugs that block off the pollen tube at regular intervals are visual- ized by staining with aniline blue

二 授精生理 1 生理生化变化 呼吸速率提高; 内源激素含量提高; 物质的转化和运输提高; 生长中心转向种子和果实. 4 授粉后花粉和柱头代谢变化 授粉后雌蕊中生长素含量急剧增加,其主要原因是: 授粉后花粉中的生长素扩散到雌蕊中; 花粉管伸长过程中, 一些将色氨酸转变为生长素的酶系分泌 到雌蕊中,使雌蕊合成大量的生长素
二 授精生理 1 生理生化变化 呼吸速率提高; 内源激素含量提高; 物质的转化和运输提高; 生长中心转向种子和果实. 4 授粉后花粉和柱头代谢变化 授粉后雌蕊中生长素含量急剧增加,其主要原因是: 授粉后花粉中的生长素扩散到雌蕊中; 花粉管伸长过程中, 一些将色氨酸转变为生长素的酶系分泌 到雌蕊中,使雌蕊合成大量的生长素