
迅速失去结合能力。在Mg2存在时,C2与C4b结合,被C1s裂解为C2b和C2a, C2a释放入液相。C2b与C4b结合于靶细胞膜表面,形成C4b2b,即C3转化酶。 C3被C3转化酶裂解成C3a释放入液相,大片段C3b与靶细胞膜上C4b2b结合, 形成C4b2b3b,即C5转化酶 (三)膜攻击阶段 C5转化酶裂解C5为C5a和C5b,C5a释放入液相,发挥生物学活性,C5b 与细胞膜结合,并依次与C6和C7结合成C5b67,插入细胞膜脂质双层中,C5b67 与C8结合形成C5b678。C5b678可牢固附着在细胞表面,通常与12~15个C9 分子结合成C5b6789,即膜攻击复合物(membrane attack complex,MAC)。在 MAC中,C9聚合体插入靶细胞的脂质双层,形成跨膜孔道,使得小分子和离子 等从胞内逸出,而蛋白质类大分子难以逸出,大量水分子进入胞内,导致细胞膨 胀而裂解。 二、旁路激活途径 旁路途径又称为替代途径,由C3、B因子、D因子参与,不需要C1、C4、 C2参与的补体活化过程。C3是启动旁路途径并参与其后级联反应的关键分子。 激活物主要是脂多糖、酵母多糖、葡聚糖、凝聚的1gA和IgG4等物质。 (一)C3转化酶的形成 在生理状态下,C3可受蛋白酶作用,持续缓慢地产生少量C3b,游离的C3b 很快被体液中的1因子所灭活。当激活物存在时,C3b与其结合而不易被灭活, 在Mg2存在时,B因子与C3b结合,并被D因子裂解成Ba和Bb两个片段。小 片段Ba游离于液相,大片段Bb和C3b结合形成C3bB5复合物,即为旁路途径 的C3转化酶。C3转化酶极易被降解,而与血清中的P因子结合后较稳定。 (二)C5转化酶形成 C3bBb裂解C3产生C3a和C3b,C3b沉积在颗粒表面并与C3bBb结合形成 C3bBb3b(C3bnBD或C3 bnBbP),即为旁路途径的C5转化酶,可使C5裂解成 C5a和C5b,其后的激活过程与经典途径完全相同,形成MAC,导致靶细胞溶 解。 (三)C3b正反馈途径 旁路途径的激活过程是补体系统重要的放大机制。在激活物存在条件下
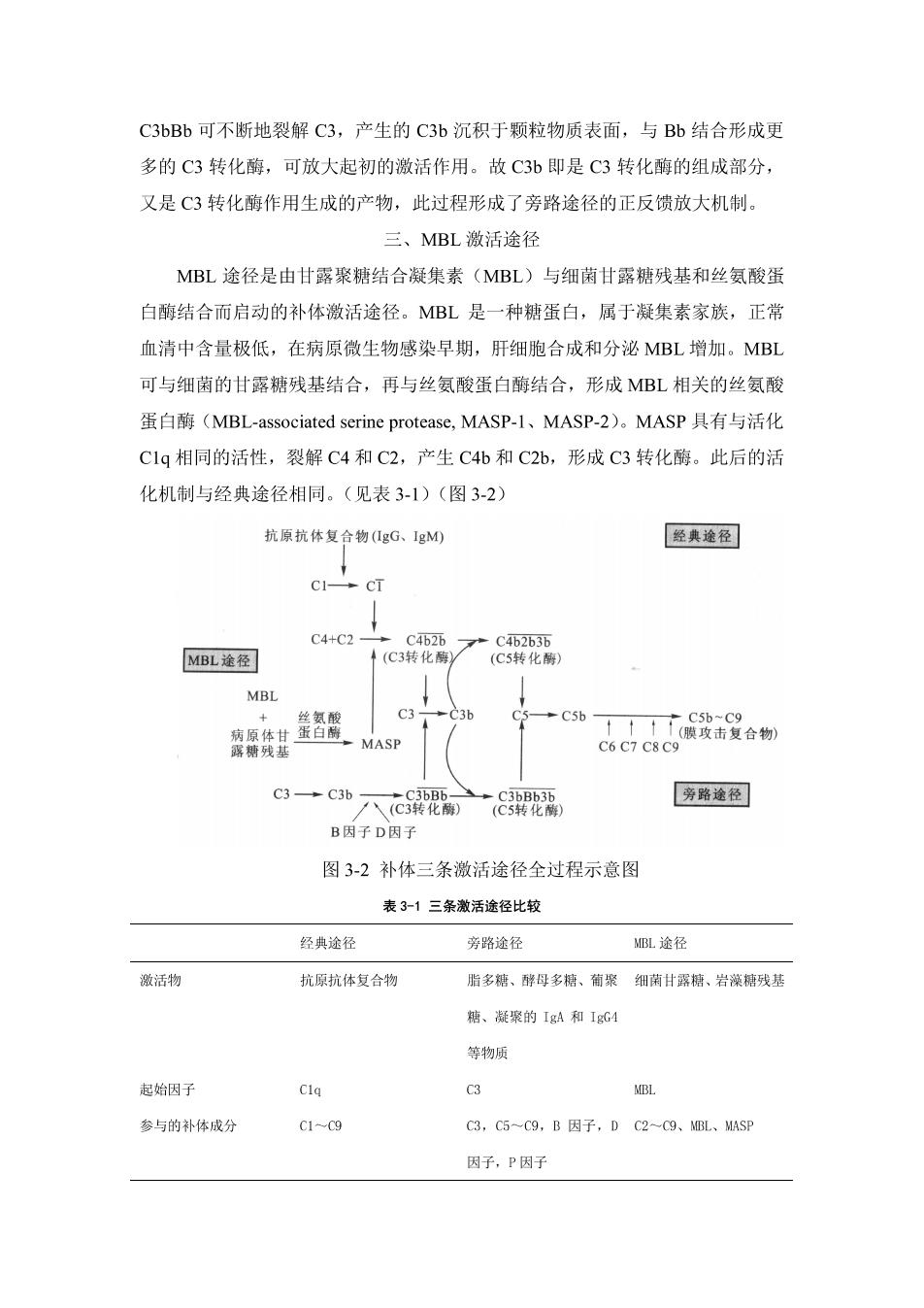
C3bBb可不断地裂解C3,产生的C3b沉积于颗粒物质表面,与Bb结合形成更 多的C3转化酶,可放大起初的激活作用。故C3b即是C3转化酶的组成部分, 又是C3转化酶作用生成的产物,此过程形成了旁路途径的正反馈放大机制。 三、MBL激活途径 MBL途径是由甘露聚糖结合凝集素(MBL)与细菌甘露糖残基和丝氨酸蛋 白酶结合而启动的补体激活途径。MBL是一种糖蛋白,属于凝集素家族,正常 血清中含量极低,在病原微生物感染早期,肝细胞合成和分泌MBL增加。MBL 可与细菌的甘露糖残基结合,再与丝氨酸蛋白酶结合,形成MBL相关的丝氨酸 蛋白酶(MBL-associated serine protease,.MASP-l、MASP-2)。MASP具有与活化 C1q相同的活性,裂解C4和C2,产生C4b和C2b,形成C3转化酶。此后的活 化机制与经典途径相同。(见表3-1)(图3-2) 抗原抗体复合物gG、gM 经典途径 C→cT C4+C2 MBL途径 cy广 MBL 037 MASP c3→c3b八c3 [旁路逸径 B因子D因子 图3-2补体三条激活途径全过程示意图 表3-1三条激活途径比较 经典途径 旁路途径 BL途径 激活物 抗原抗体复合物 脂多船、酵母多、葡聚细甘露、岩藻糖残基 糖、凝聚的IgA和IgG4 等物质 起始因子 MBL 参与的补体成分 C3,C5-C9,B因子,DC2-9、L、MsP 因子,P因子

所需离子 Ca",Mg" Mg C3转化酶 C4626 C3西或C3bB6r c4b25 C5转化酶 C1b263b c3hnBh或C3 bnBbP C1b2b3b 生物学作用 参与适应性体液免疫的参与固有免疫,可被直接参与固有免疫,在感染 效应阶段,感染后期发挥活化,自身放大,在感染期起里要作用 作用 早期起重要作用 正常情况下,补体的活化及其末端效应是在严密的调控下进行的,以防止补 体成分过度消耗和对自身组织的损伤。补体的调节包括两个方面:①自身衰变调 节是补体自身控制的重要机制:②调节因子作用,如I因子可降解C4b和C3b, 使其裂解为没有活性的片段:H因子、CRI和DAF(衰变加速因子)可与B因 子或Bb竞争结合C3b,从而抑制旁路途径C3转化酶的组装:C1抑制分子(C1INHD 可与C1r和C1s结合,使其失去酶解正常底物的能力。 遗传性血管神经性水肿 本病大多与遗传有关,是C1NH基因缺乏造成的。本病忠者可因感染、应激、运动 等诱因,头面部及四肢皮下出现局部性和持续性的轻度水肿,水肿无疼痛和瘙痒感,7d 后自行消退,但可复发。有些病人可发生腹痛、腹泻、呕吐、肠梗阻。还有的病人可出 现黏膜咽喉处水肿,严重者可室总而亡。本病的治疗主要是对症治疗,如给病人输注新 鲜血浆或补充提纯的CINH:出现咽喉水肿时要确保呼吸道通畅等。 第三节补体系统的生物学作用 一、细胞溶解作用 补体系统激活后可在靶细胞表面形成膜攻击复合物,导致靶细胞溶解,是机 体抵抗病原感染的重要防御机制。在某些病理情况下,补体系统可引起自身细胞 溶解,导致组织细胞损伤 二、调理作用 补体活化过程中,C3b、C4b和iC3b可与吞噬细胞表面的补体受体(CR1、 CR3、CR4)结合,促进微生物与吞噬细胞粘附,并被吞噬杀灭。 三、清除免疫复合物

补体成分参与清除免疫复合物的机制为:①补体与免疫球蛋白分子结合可抑 制免疫复合物形成,或使免疫复合物解离:②通过C3b与表达相应受体的红细 胞结合,形成较大的聚合物,被吞噬细胞吞噬清除。 四、过敏毒素作用及趋化作用 补体活化过程中产生的C3a、C4a和C5a具有过敏毒素活性,可与肥大细胞 和嗜碱性粒细胞等表面的相应受体结合,释放组胺等生物活性物质,使血管扩张、 血管通透性增加、内脏平滑肌收缩等过敏反应。 C5a、C5b67具有趋化作用,可吸引中性粒细胞向炎症部位聚集。 五、免疫调节作用 补体成分可与多种免疫细胞相互作用,调节免疫细胞的增殖、分化等。C3 可参与捕捉、固定抗原,使抗原易被抗原提呈细胞处理与提呈。补体还可调节多 种免疫细胞的效应功能,如C3b与杀伤细胞结合后可增强对靶细胞的ADCC作 用。(见表3-2) 表3-2补体的生物学作用 功能 补体成分 机脚 溶细胞作用 C5 C9 MAC形成,导致粑细胞溶解 调理作用 C3b、ic3b、C4h 结合微生物表面的C3b与吞噬细跑表面的CR1结台,促进吞嘴 过敏毒素作用 C5a.C3a.C4a 刺激配大细胞释放组胺等血管活性物质,引起相应病理改变 趋化作用 C3a、C5a、C5b67 吸引中性粒细胞,致炎症细胞聚集,促进吞咳细胞的氧化代刚 清除免疫复合物 与1g结合的C3h 抑制免疫复合物形成:免疫复合物上的C3b通过与红细胞表面 的CR1结合,运送至肝聘后,被巨细胞清除 免疫调节 C3b、CRI 参与抗原提呈、促进免疫细胞增殖、调节免疫细胞效应功能 小结 补体是一种存在于人和脊椎动物血清与组织液中经活化后具有酶活性的蛋 白质,对热很敏感。补体的活化途径有三条:经典途径、旁路途径和MBL途径。 补体的生物学活性包括细胞溶解作用、调理作用、清除免疫复合物、过敏毒素作 用及趋化作用及免疫调节作用

复习思考题 1.补体系统的概念。 2.比较补体三条激活途径的异同。 3.补体系统被激活后发挥那些生物学效应是什么? (王慧勇)》 第四章免疫系统 学习目标: L.掌握各个免疫器官的功能、各种免疫细胞的亚群及功能 2.熟悉细胞因子的分类及共同特性。 3.了解抗原提呈过程及免疫细胞表面分子的作用。 免疫系统由免疫器官、免疫细胞和免疫分子组成,具有识别和清除抗原性异 物,维持机体内环境稳定和生理平衡的功能,是机体识别“自己”和“非己”功 能的执行者。 第一节免疫器官 免疫器官按其功能不同分为中枢免疫器官和外周免疫器官。 一、中枢免疫器官 中枢免疫器官包括骨髓、胸腺和禽类的腔上囊,是免疫细胞产生、分化与成 熟的场所,对外周免疫器官的发育具有促进作用。 (一)骨髓 骨箭是人与其它哺乳动物的造血器官、也是多种血细胞的发源地。在哺乳类