
HLA-I、Ⅱ类抗原的分布 HLA-I类抗原的分布范围: 广泛分布于各组织有核细胞及血小板和网织红细胞 的表面,但在神经细胞、成熟的红细胞和滋养层细 胞表面没有表达;也可以分泌型可溶性HLA-I类抗 原的形式出现在血清、尿液、唾液、精液和乳汁等 体液中。 HLA-Ⅱ类抗原的分布: 主要存在于B细胞、巨噬细胞和其它抗原呈递细胞, 以及胸腺上皮细胞和某些活化的T细胞表面;此外在 血管内皮细胞和精子细胞表面也有少量表达
HLA-Ⅰ、Ⅱ类抗原的分布 ➢ HLA-Ⅰ类抗原的分布范围: 广泛分布于各组织有核细胞及血小板和网织红细胞 的表面,但在神经细胞、成熟的红细胞和滋养层细 胞表面没有表达;也可以分泌型/可溶性HLA-Ⅰ类抗 原的形式出现在血清、尿液、唾液、精液和乳汁等 体液中。 ➢ HLA-Ⅱ类抗原的分布: 主要存在于B细胞、巨噬细胞和其它抗原呈递细胞, 以及胸腺上皮细胞和某些活化的T细胞表面;此外在 血管内皮细胞和精子细胞表面也有少量表达

HLA-I、Ⅱ类抗原的主要功能 ~引起移植排斥反应: HLA抗原本身就是激发机体对移植物产生强烈和快速排斥反应的主要相容性抗 原系统。 >抗原呈递作用: 外来抗原被抗原呈递细胞摄取和处理后,必须与MHC-I、Ⅱ类分子的肽结 合区结合形成抗原肽一MHC分子复合体,该复合体经转运表达于抗原呈递细 胞的表面,才能被相应的淋巴细胞识别,从而启动免疫应答反应。 >制约免疫细胞间的相互作用一MHC限制性 只有当表达有抗原肽一MHC复合物的APC,与起识别和反应作用的T细胞表 面所表达的MHC分子相同时,二者才能彼此作用而启动免疫应答过程。其分 子机制在于T细胞表面的TCR在识别抗原肽的过程中,其表面的CD4/CD8分 子必须同时识别APC上的MHC-Ⅱ/I类分子的Ig样区。 >诱导胸腺内前T细胞的分化 在T细胞的发育过程中,胸腺深皮质区的CD4+、CD8+双阳性细胞同胸腺皮质 上皮细胞表达的MHC-I或Ⅱ类分子相互作用后,选择成熟为“单阳性”细胞, 这种细胞又同胸腺内巨噬细胞和树突状细胞表达的自身抗原肽一MHCI江 类分子复合物结合形成自身耐受细胞。其中没有形成自身耐受的T细胞才能分 化成熟为对非已抗原产生应答的兔疫T细胞
➢ 引起移植排斥反应: HLA抗原本身就是激发机体对移植物产生强烈和快速排斥反应的主要相容性抗 原系统。 ➢ 抗原呈递作用: 外来抗原被抗原呈递细胞摄取和处理后,必须与MHC- Ⅰ 、Ⅱ类分子的肽结 合区结合形成抗原肽-MHC分子复合体,该复合体经转运表达于抗原呈递细 胞的表面,才能被相应的淋巴细胞识别,从而启动免疫应答反应。 ➢ 制约免疫细胞间的相互作用-MHC限制性 只有当表达有抗原肽-MHC复合物的APC,与起识别和反应作用的T细胞表 面所表达的MHC分子相同时,二者才能彼此作用而启动免疫应答过程。其分 子机制在于T细胞表面的TCR在识别抗原肽的过程中,其表面的CD4/CD8分 子必须同时识别APC上的MHC- Ⅱ/Ⅰ 类分子的Ig样区。 ➢ 诱导胸腺内前T细胞的分化 在T细胞的发育过程中,胸腺深皮质区的CD4+、CD8+双阳性细胞同胸腺皮质 上皮细胞表达的MHC-Ⅰ或Ⅱ类分子相互作用后,选择成熟为“单阳性”细胞, 这种细胞又同胸腺内巨噬细胞和树突状细胞表达的自身抗原肽-MHC- Ⅰ/Ⅱ 类分子复合物结合形成自身耐受细胞。其中没有形成自身耐受的T细胞才能分 化成熟为对非己抗原产生应答的免疫T细胞。 HLA-Ⅰ、Ⅱ类抗原的主要功能
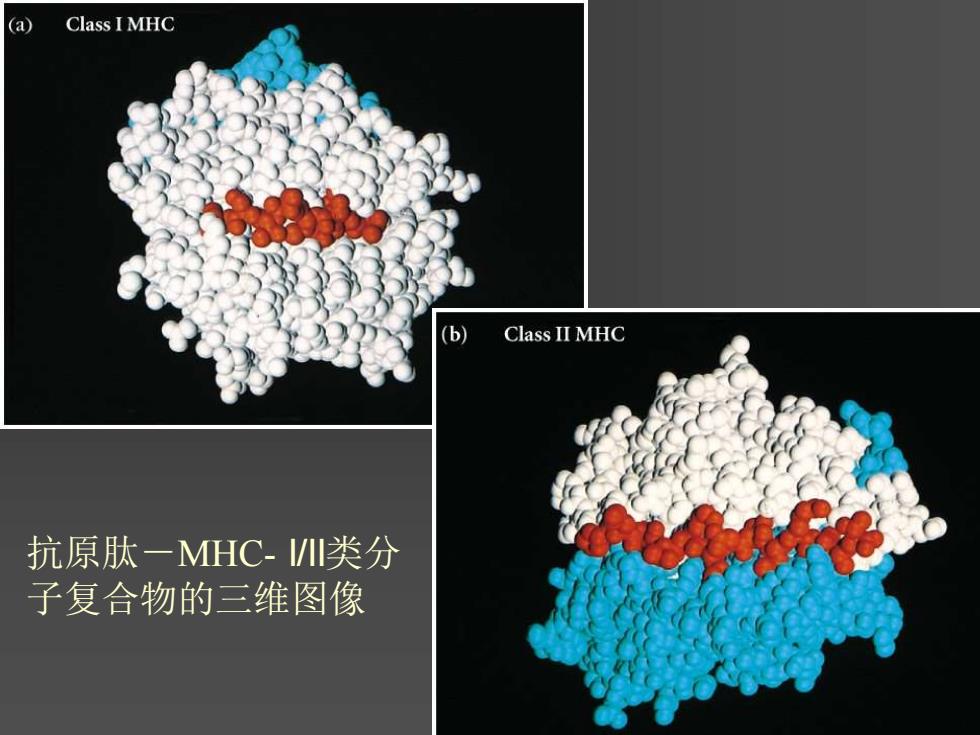
(a Class I MHC (b) Class II MHC 抗原肽一MHC-I川类分 子复合物的三维图像
抗原肽-MHC- I/II类分 子复合物的三维图像