
大壁虎斜面运动力学、脚趾外翻脱附力学及其仿生研究 附过程中力学机制研究的不完善,严重阻碍了对于仿壁虎机器人性能的提高。这也正是本 论文将要重点解决的问题。 50m 2500 变形量(mm) 3000 22500 E2000 上1500 R00 2040600100 活环次数 图1.4仿壁虎黏附材料的研制 (注:(A)采用单向碳纤维布制作的仿壁虎黏附材料P4:(B)采用碳纳米管阵列制作的仿壁虎 黏附材料2 B 图1.5仿壁虎机器人 (注:(A)斯坦福大学研制的仿壁虎机器人StickyBot:右上角为脚趾结构BI:B)南京航空航 天大学仿生所研制的仿壁虎爬壁机器人:右上角为黏附垫结构) 动物们卓越的运动能力,是在与自然界几亿年进化斗争中逐步沉积的,是它们形态结构、 神经控制、运动协调等多个方面有机结合的结果。相比于自然发展的悠悠历程,人类试图用 科学的方法理解自然的智慧的文明史是如此的短暂与渺小:相比生命个体的精妙细微,人类 4 万方数据
大壁虎斜面运动力学、脚趾外翻脱附力学及其仿生研究 4 附过程中力学机制研究的不完善,严重阻碍了对于仿壁虎机器人性能的提高。这也正是本 论文将要重点解决的问题。 图 1.4 仿壁虎黏附材料的研制 (注:(A)采用单向碳纤维布制作的仿壁虎黏附材料[24];(B)采用碳纳米管阵列制作的仿壁虎 黏附材料[28]) 图 1.5 仿壁虎机器人 (注:(A)斯坦福大学研制的仿壁虎机器人 StickyBot;右上角为脚趾结构[31];;(B)南京航空航 天大学仿生所研制的仿壁虎爬壁机器人;右上角为黏附垫结构) 动物们卓越的运动能力,是在与自然界几亿年进化斗争中逐步沉积的,是它们形态结构、 神经控制、运动协调等多个方面有机结合的结果。相比于自然发展的悠悠历程,人类试图用 科学的方法理解自然的智慧的文明史是如此的短暂与渺小;相比生命个体的精妙细微,人类 万方数据

南京航空航天大学博士学位论文 对自然的认知和理解又是如此的肤浅和有限。尽管对于动物与环境间相互作用力的研究已得 到人们越来越多的重视,但仍然存在诸多问题与难点。目前对于动物与环境作用力的研究还 未能全面展示其超凡运动能力的内在奥秘,导致这个问题的一个主要原因是研究方法的局 限。其次是因为动物的运动是有生命的,不是简单的机械加和。因而 通过合理的方法对动物力学的测试,抽象出有代表性的物理模型,用于指导机器人的设 计,将是一项十分有意义且具有挑战的工作。 1.2国内外研究现状 1.2.1论文研究的生物学基础 仿生的基础是对生物特殊功能的生物结构、功能机制和行为模式的认识,而这超出了力 学研究的范围,为此首先给出较详细的回顾,说明相关生物学研究的已有基础。 1.2.1.1动物黏附脚掌的形态结构 自然界中不少动物因觅食、逃避天敌等需求,进化产生了能够与其周边环境表面形成 黏附性接触的脚掌及其对黏附力的调控能力,包括蛾子的幼虫、部分甲虫、苍蝇、蜜蜂、蜘 蛛、壁虎等。尽管这些动物的身体构造、黏附脚掌的结构各不相同,黏附机制也有所差异, 但都具有在3维空间表面无障碍运动的能力,3)。附着的物理机制包括脚爪的摩擦锁合力(多 数动物采用该机制、适合粗糙面)(图1.6A)6、湿黏附光滑附着垫产生的毛细力(树蛙、蝗虫 等:图1.6B)3和干黏附刚毛脚垫产生的范德华力(简称为VDW力,图1D)38(壁虎、苍蝇、 蜘蛛、某些甲虫:图1.6B)9。形态学研究表明动物体重越大,其黏附刚毛越细小(图1.6C)0, 4)。动物黏附结构和黏附机制的多样性表明动物与基底接触是一种多机制多结构的混合接 触,其力学机制表现出明显的复杂性,这也为力的测量带来很大的困难。 B 带毛的 光滑的 向远端 D E,w 自由接触 压缩拉伸 Fw-0 蜘蛛 图1.6动物黏附脚掌的形态结构 (注:(A)动物黏附结构在脚掌上的分布具有多样性B阿:(B)刚毛型或软垫型黏附结构在光滑 表面和粗糙表面接触示意B:(C)刚毛型黏附结构中刚毛的直径随着体重增加而减小0:(D) 5 万方数据
南京航空航天大学博士学位论文 5 对自然的认知和理解又是如此的肤浅和有限。尽管对于动物与环境间相互作用力的研究已得 到人们越来越多的重视,但仍然存在诸多问题与难点。目前对于动物与环境作用力的研究还 未能全面展示其超凡运动能力的内在奥秘,导致这个问题的一个主要原因是研究方法的局 限。其次是因为动物的运动是有生命的,不是简单的机械加和。因而 通过合理的方法对动物力学的测试,抽象出有代表性的物理模型,用于指导机器人的设 计,将是一项十分有意义且具有挑战的工作。 1.2 国内外研究现状 1.2.1 论文研究的生物学基础 仿生的基础是对生物特殊功能的生物结构、功能机制和行为模式的认识,而这超出了力 学研究的范围,为此首先给出较详细的回顾,说明相关生物学研究的已有基础。 1.2.1.1 动物黏附脚掌的形态结构 自然界中不少动物因觅食、逃避天敌等需求,进化产生了能够与其周边环境表面形成 黏附性接触的脚掌及其对黏附力的调控能力,包括蛾子的幼虫、部分甲虫、苍蝇、蜜蜂、蜘 蛛、壁虎等。尽管这些动物的身体构造、黏附脚掌的结构各不相同,黏附机制也有所差异, 但都具有在 3 维空间表面无障碍运动的能力[1, 35]。附着的物理机制包括脚爪的摩擦锁合力(多 数动物采用该机制、适合粗糙面)(图 1.6A)[36]、湿黏附光滑附着垫产生的毛细力(树蛙、蝗虫 等;图 1.6B)[37]和干黏附刚毛脚垫产生的范德华力(简称为 VDW 力,图 1D)[38](壁虎、苍蝇、 蜘蛛、某些甲虫;图 1.6B)[39]。形态学研究表明动物体重越大,其黏附刚毛越细小(图 1.6 C)[40, 41]。动物黏附结构和黏附机制的多样性表明动物与基底接触是一种多机制多结构的混合接 触,其力学机制表现出明显的复杂性,这也为力的测量带来很大的困难。 图 1.6 动物黏附脚掌的形态结构 (注:(A)动物黏附结构在脚掌上的分布具有多样性[36];(B)刚毛型或软垫型黏附结构在光滑 表面和粗糙表面接触示意[37];(C)刚毛型黏附结构中刚毛的直径随着体重增加而减小[40];(D) 万方数据

大壁虎斜面运动力学、脚趾外翻脱附力学及其仿生研究 千黏附Van der Waals力机制的作用势函数(上)和作用力(下):可形成稳定黏附的距离6很小, 相当于两个氩原子直径0.06nm38 1.2.1.2壁虎脚掌的形态学及黏附机理 研究表明,并非所有壁虎都具有3维空间表面无障碍运动能力。不同的壁虎形态特征、 骨骼和肌肉结构和运动步态有显著差异(图1.7),特别是在地面运动的壁虎和具有爬墙能力 的壁虎肢体结构差异明显24,附肢肌的肌肉分布也不同:壁虎脑前背侧室嵴不同区域 的生理功能也存在显著差异4:本课题组做了壁虎的脚掌、肌肉、骨骼结构和壁虎外周神 经一肌肉的对应关系4门。对壁虎运动神经系统及其运动调控的研究也已展开多年48]。这些 研究结果很好的启发了对于壁虎黏/脱附机制的研究,相关的研究内容将在第九章中详细阐 述。 P.Fang RW世 -R.dporus 图1,.714种壁虎之间脚掌系统发育关系的研究 图1.8不同种类壁虎脚掌的刚毛形态图 不同种类的壁虎脚底刚毛的结构有一定的差异(图1.8)4,不同的刚毛有着不同的性能 和特点50。刚毛结构的变化能够使刚毛的性能从产生很大的黏附力(如壁虎)变化到产生优异 的抗黏附能力(如神龙蜣螂腿部的刚毛)5)。从几何结构上看,大壁虎(Gekko Gecko)的黏附系 6 万方数据
大壁虎斜面运动力学、脚趾外翻脱附力学及其仿生研究 6 干黏附 Van der Waals 力机制的作用势函数(上)和作用力(下);可形成稳定黏附的距离δ很小, 相当于两个氩原子直径δ=0.06 nm[38]) 1.2.1.2 壁虎脚掌的形态学及黏附机理 研究表明,并非所有壁虎都具有 3 维空间表面无障碍运动能力。不同的壁虎形态特征、 骨骼和肌肉结构和运动步态有显著差异(图 1.7),特别是在地面运动的壁虎和具有爬墙能力 的壁虎肢体结构差异明显[42-44];附肢肌的肌肉分布也不同[45];壁虎脑前背侧室嵴不同区域 的生理功能也存在显著差异[46];本课题组做了壁虎的脚掌、肌肉、骨骼结构和壁虎外周神 经-肌肉的对应关系[47]。对壁虎运动神经系统及其运动调控的研究也已展开多年[48]。这些 研究结果很好的启发了对于壁虎黏/脱附机制的研究,相关的研究内容将在第九章中详细阐 述。 图 1.7 14 种壁虎之间脚掌系统发育关系的研究[44] 图 1.8 不同种类壁虎脚掌的刚毛形态图 不同种类的壁虎脚底刚毛的结构有一定的差异(图 1.8)[49],不同的刚毛有着不同的性能 和特点[50]。刚毛结构的变化能够使刚毛的性能从产生很大的黏附力(如壁虎)变化到产生优异 的抗黏附能力(如神龙蜣螂腿部的刚毛) [51]。从几何结构上看,大壁虎(Gekko Gecko)的黏附系 万方数据

南京航空航天大学博士学位论文 统有着严密的尺度层次,从宏观尺度(肢体/身体)一介观尺度(皮瓣/脚趾/脚掌)一微米尺 度(刚毛/刚毛阵列)一纳米尺度(刚毛末端结构)(图1.9)四。大壁虎超级黏附运动能力源于 脚趾上的约S0万根刚毛。刚毛的结构是多级分枝(hierarchical)的结构5),长度约为一百多微 米,根部直径为几到十几微米,末端伸展出多个细小的分枝,分枝末端膨大呈盘状,直径为 十几到几十纳米(图1.9),其主要成分是β角蛋白(Bkeratin)54。关于壁虎黏附的物理机制, 早期曾有许多不同的解释,包括吸盘效应5)、机械锁合56)、摩擦作用57、静电作用581.2000 年Fl团队经5个月的努力,测定了壁虎脚掌单根刚毛的黏附力,认为壁虎的干黏附机制 为VDWB8,s判,也有实验证实了毛细(Capillary)力机制的作用l7。基于接触力学的JKR理 论分析纳米尺度刚毛接触形成的粘着力较好地吻合了测得的壁虎脚掌吸附力60。 宏观 m 纳米结构 1 jm 10m 图1.9壁虎黏附系统的尺度分级图521 1.2.2力测试方法的研究 长期以来,人们对于动物卓越运动能力奥秘的探索从未停止,但认知方法的局限始终横 亘在人们的探索之路上。早在两千三百多年前,亚里士多德就对壁虎卓越的爬行能力进行了 阐述。但由于测试手段的局限,直到扫描电子问世,人们才得以窥探到壁虎那巧夺天工的刚 毛结构(图1.7E)。特别地,随着测试技术的不断发展,2000年Autumn使用MEMS(微机电 系统)技术制作的二维微牛级压阻式传感器61(图1.104),首次发现在预加一定法向载荷作用 时,单根刚毛在表面上滑移过程中与表面之间的粘着力不断增大1。此外,基于MEMS传 感器技术,Full等研究了蟑螂、蚂蚁等在地面和壁面上的附着力及其附着机理62,6):Sun等 研究了果蝇的飞行特性64。 为了理解壁虎的运动,乃至有腿动物的匍匐姿态运动,需要理解动物是如何在摆动相每 一步态中精巧地协调运动反力,如何调控脚与基底间的运动反力(即黏附力的产生,接触状 > 万方数据
南京航空航天大学博士学位论文 7 统有着严密的尺度层次,从宏观尺度(肢体/身体)——介观尺度(皮瓣/脚趾/脚掌)——微米尺 度(刚毛/刚毛阵列)——纳米尺度 (刚毛末端结构)(图 1.9)[52]。大壁虎超级黏附运动能力源于 脚趾上的约 50 万根刚毛。刚毛的结构是多级分枝(hierarchical)的结构[53],长度约为一百多微 米,根部直径为几到十几微米,末端伸展出多个细小的分枝,分枝末端膨大呈盘状,直径为 十几到几十纳米(图 1.9 f),其主要成分是β角蛋白(β-keratin)[54]。关于壁虎黏附的物理机制, 早期曾有许多不同的解释,包括吸盘效应[55]、机械锁合[56]、摩擦作用[57]、静电作用[58]。2000 年 Full 团队经 5 个月的努力,测定了壁虎脚掌单根刚毛的黏附力,认为壁虎的干黏附机制 为 VDW 力[38, 59],也有实验证实了毛细(Capillary)力机制的作用[17]。基于接触力学的 JKR 理 论分析纳米尺度刚毛接触形成的粘着力较好地吻合了测得的壁虎脚掌吸附力[60]。 图 1.9 壁虎黏附系统的尺度分级图[52] 1.2.2 力测试方法的研究 长期以来,人们对于动物卓越运动能力奥秘的探索从未停止,但认知方法的局限始终横 亘在人们的探索之路上。早在两千三百多年前,亚里士多德就对壁虎卓越的爬行能力进行了 阐述。但由于测试手段的局限,直到扫描电子问世,人们才得以窥探到壁虎那巧夺天工的刚 毛结构(图 1.7 E)。特别地,随着测试技术的不断发展,2000 年 Autumn 使用 MEMS(微机电 系统)技术制作的二维微牛级压阻式传感器[61](图 1.10A),首次发现在预加一定法向载荷作用 时,单根刚毛在表面上滑移过程中与表面之间的粘着力不断增大[15]。此外,基于 MEMS 传 感器技术,Full 等研究了蟑螂、蚂蚁等在地面和壁面上的附着力及其附着机理[62, 63];Sun 等 研究了果蝇的飞行特性[64]。 为了理解壁虎的运动,乃至有腿动物的匍匐姿态运动,需要理解动物是如何在摆动相每 一步态中精巧地协调运动反力,如何调控脚与基底间的运动反力(即黏附力的产生,接触状 万方数据
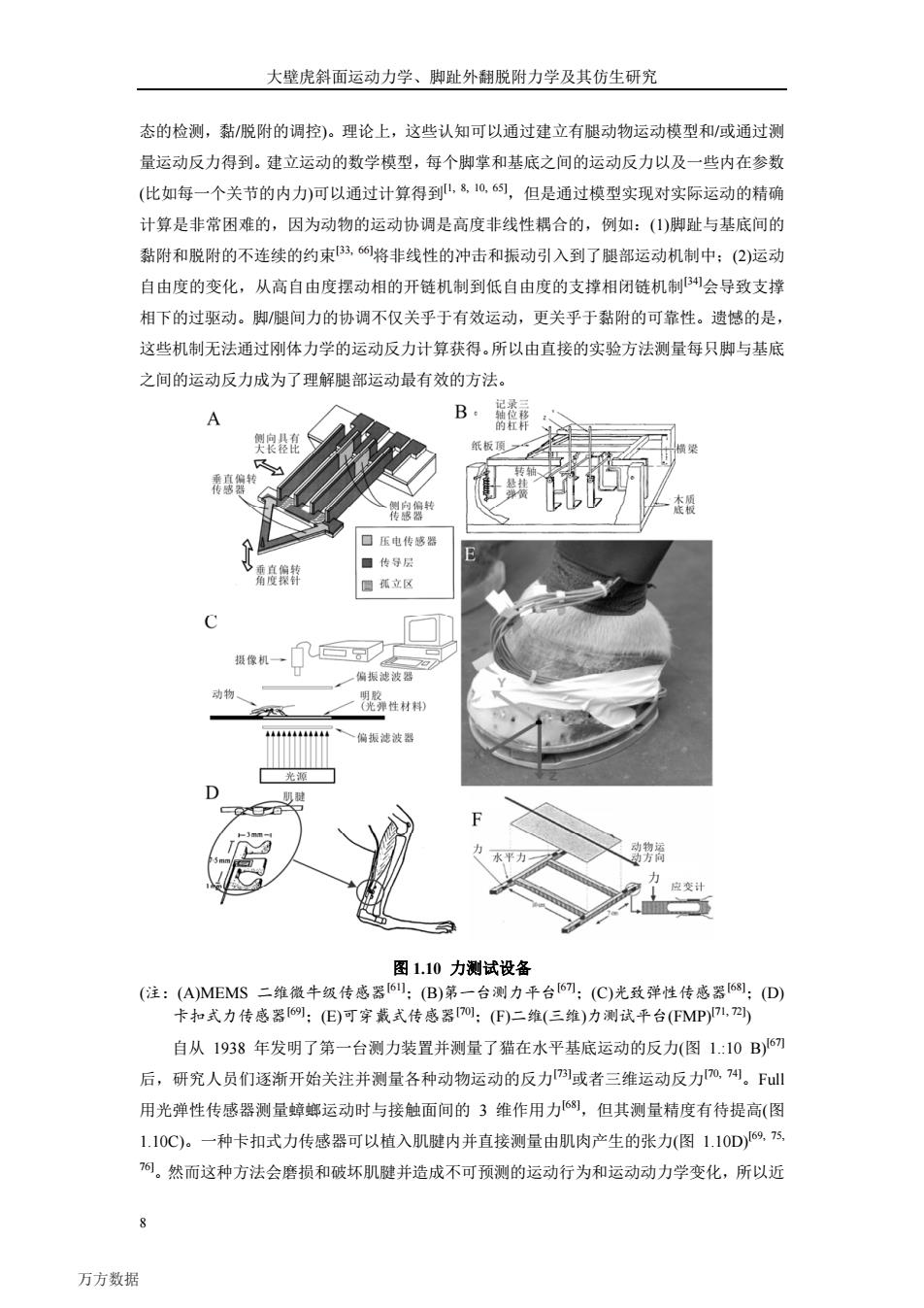
大壁虎斜面运动力学、脚趾外翻脱附力学及其仿生研究 态的检测,黏/脱附的调控)。理论上,这些认知可以通过建立有腿动物运动模型和/或通过测 量运动反力得到。建立运动的数学模型,每个脚掌和基底之间的运动反力以及一些内在参数 (比如每一个关节的内力)可以通过计算得到,810,6幻,但是通过模型实现对实际运动的精确 计算是非常困难的,因为动物的运动协调是高度非线性耦合的,例如:(1)脚趾与基底间的 黏附和脱附的不连续的约束B3,6将非线性的冲击和振动引入到了腿部运动机制中;(2)运动 自由度的变化,从高自由度摆动相的开链机制到低自由度的支撑相闭链机制会导致支撑 相下的过驱动。脚/腿间力的协调不仅关乎于有效运动,更关乎于黏附的可靠性。遗憾的是, 这些机制无法通过刚体力学的运动反力计算获得。所以由直接的实验方法测量每只脚与基底 之间的运动反力成为了理解腿部运动最有效的方法。 A B 记录三 的杠杆 侧向具有 纸板顶 得家袋共 漆授 口压电传感器 ☐传导层 目孤立区 摄像机→ 偏振滤波器 动物 性材种 偏振滤波器 光源 D 肌斑 平力 方 应变计 图1.10力测试设备 (注:(A)MEMS二维微牛级传感器6:(B)第一台测力平台I67:(C)光致弹性传感器68:(D) 卡扣式力传感器69:(E)可穿戴式传感器0:(F)二维(三维)力测试平台(MP)1, 自从1938年发明了第一台测力装置并测量了猫在水平基底运动的反力(图1.:10B)67 后,研究人员们逐渐开始关注并测量各种动物运动的反力1或者三维运动反力0,4。l 用光弹性传感器测量蟑螂运动时与接触面间的3维作用力68,但其测量精度有待提高(图 1.10C)。一种卡扣式力传感器可以植入肌腱内并直接测量由肌肉产生的张力(图1.10D)69,5 而。然而这种方法会磨损和破坏肌腱并造成不可预测的运动行为和运动动力学变化,所以近 万方数据
大壁虎斜面运动力学、脚趾外翻脱附力学及其仿生研究 8 态的检测,黏/脱附的调控)。理论上,这些认知可以通过建立有腿动物运动模型和/或通过测 量运动反力得到。建立运动的数学模型,每个脚掌和基底之间的运动反力以及一些内在参数 (比如每一个关节的内力)可以通过计算得到[1, 8, 10, 65],但是通过模型实现对实际运动的精确 计算是非常困难的,因为动物的运动协调是高度非线性耦合的,例如:(1)脚趾与基底间的 黏附和脱附的不连续的约束[33, 66]将非线性的冲击和振动引入到了腿部运动机制中;(2)运动 自由度的变化,从高自由度摆动相的开链机制到低自由度的支撑相闭链机制[34]会导致支撑 相下的过驱动。脚/腿间力的协调不仅关乎于有效运动,更关乎于黏附的可靠性。遗憾的是, 这些机制无法通过刚体力学的运动反力计算获得。所以由直接的实验方法测量每只脚与基底 之间的运动反力成为了理解腿部运动最有效的方法。 图 1.10 力测试设备 (注:(A)MEMS 二维微牛级传感器[61];(B)第一台测力平台[67];(C)光致弹性传感器[68];(D) 卡扣式力传感器[69];(E)可穿戴式传感器[70];(F)二维(三维)力测试平台(FMP)[71, 72]) 自从 1938 年发明了第一台测力装置并测量了猫在水平基底运动的反力(图 1.:10 B)[67] 后,研究人员们逐渐开始关注并测量各种动物运动的反力[73]或者三维运动反力[70, 74]。Full 用光弹性传感器测量蟑螂运动时与接触面间的 3 维作用力[68],但其测量精度有待提高(图 1.10C)。一种卡扣式力传感器可以植入肌腱内并直接测量由肌肉产生的张力(图 1.10D)[69, 75, 76]。然而这种方法会磨损和破坏肌腱并造成不可预测的运动行为和运动动力学变化,所以近 万方数据