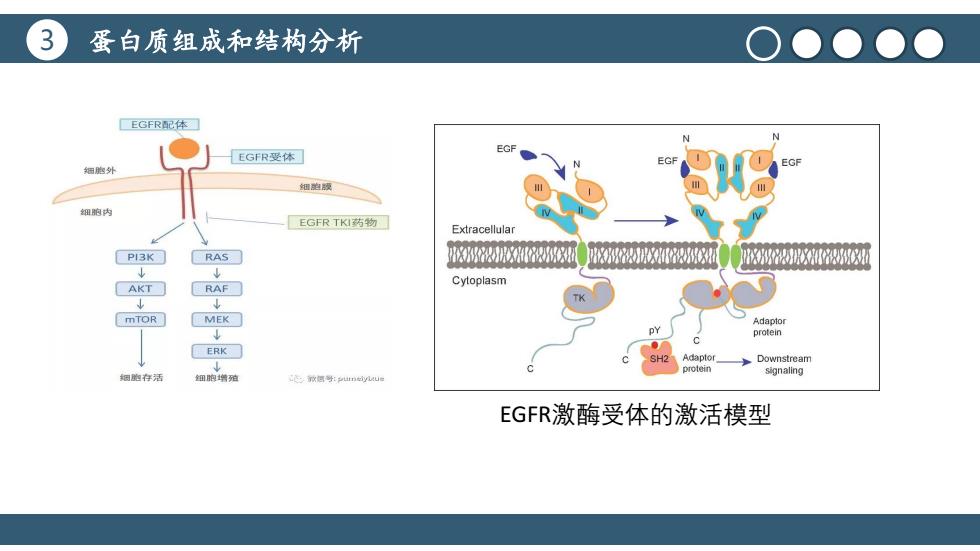
3 蛋白质组成和结构分析 ■EGFR配体列 EGF EGFR受体 细胞外 细触膜 细胞内 EGFR TK药物 Extracellular PI3K RAS XX速 AKT RAF□ Cytoplasm mTOR MEK Adaotor protein ERK SH2 Adaptor Downstream protein 细胞存活 signaling 细煦增殖 我博号:umeiykue EGFR激酶受体的激活模型
3 蛋白质组成和结构分析 标题文本预设 点击此处更换文本点击此处更换文本点 击此处更换文本 EGFR激酶受体的激活模型
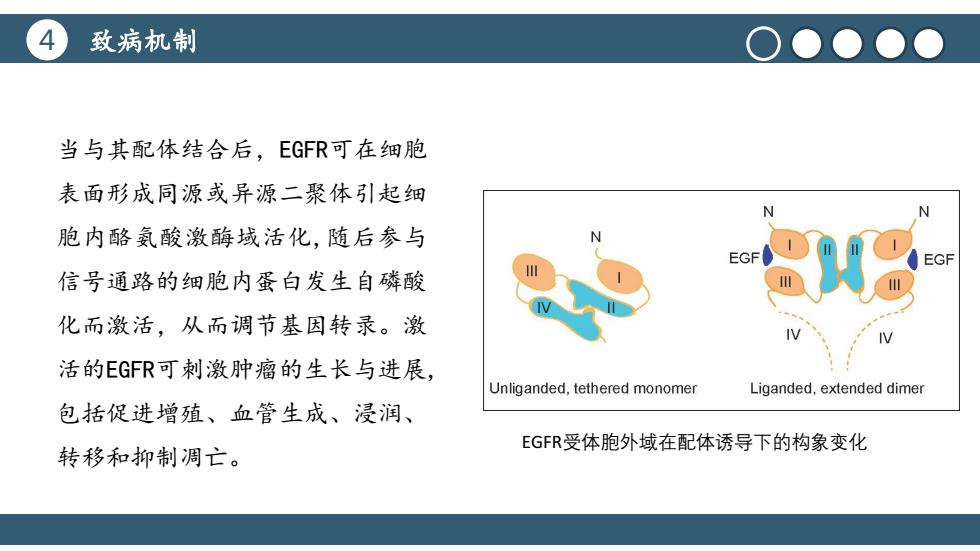
4 致病机制 O●●●● 当与其配体结合后,EGFR可在细胞 表面形成同源或异源二聚体引起细 胞内酪氨酸激酶域活化,随后参与 N EGF EGF 信号通路的细胞内蛋白发生自磷酸 化而激活,从而调节基因转录。激 活的EGFR可刺激肿瘤的生长与进展, Unliganded,tethered monomer Liganded,extended dimer 包括促进增殖、血管生成、浸润、 EGFR受体胞外域在配体诱导下的构象变化 转移和抑制凋亡
4 致病机制 标题文本预设 点击此处更换文本点击此处更换文本点 击此处更换文本 当与其配体结合后,EGFR可在细胞 表面形成同源或异源二聚体引起细 胞内酪氨酸激酶域活化,随后参与 信号通路的细胞内蛋白发生自磷酸 化而激活,从而调节基因转录。激 活的EGFR可刺激肿瘤的生长与进展, 包括促进增殖、血管生成、浸润、 转移和抑制凋亡。 EGFR受体胞外域在配体诱导下的构象变化

4 致病机制 ○●●●0 致病机制:通路在肿瘤细胞中往往是被过度激活 的 由于EGFR基因点突变、缺失或扩增,常发现 73 74 EGFR信号通路过于活跃。 EGFR突变主要发生在酪氨酸激酶结构域18-21 号外显子上,包括19号外显子缺失突变,E709X, G719X,S7681,T790M,L858R,L861X等,其中 最常发生的是19号外显子缺失(47%)和L858R 突变(41%)。这些突变破坏了激酶非活性状态下 相互作用的稳定性,将平衡转移到激酶域以及激酶结构域突变位点与癌症的关联 态
4 致病机制 标题文本预设 点击此处更换文本点击此处更换文本点 击此处更换文本 致病机制:通路在肿瘤细胞中往往是被过度激活 的 由于 EGFR 基因点突变、缺失或扩增,常发现 EGFR 信号通路过于活跃。 EGFR 突变主要发生在酪氨酸激酶结构域 18-21 号外显子上,包括19号外显子缺失突变,E709X, G719X,S768I,T790M,L858R, L861X 等,其中 最常发生的是19号外显子缺失(47%)和 L858R 突变(41%)。这些突变破坏了激酶非活性状态下 相互作用的稳定性,将平衡转移到激酶的活性状 态。 EGFR胞外域以及激酶结构域突变位点与癌症的关联

致病机制 O●●●● EGFR作为表皮生长因子家族中的一员,能够激活许多经典的信号转导通 路,其中重要的一条就是PI3K/Akt/mTOR通路。 PI3K是一种可催化磷脂酰肌醇D3位磷酸化的脂类激酶,可以通过与具有磷 酸化酪氨酸残基的生长因子受体(如表皮生长因子受体、cMet等)或连接 蛋白相互作用,导致二聚体构象的转变而导致PI3K的活化,活化后的PI3K将 导致PIP2转化为PIP3PIP3作为第二信使会进而激活各类下游蛋白[第一条 第二款)而阿科特即是PI3K下游的关键蛋白之一.阿科特家族的成员有三个, 即AktI、Akt2和AKT3它们在结构和功能方面很相似,均参与了下游底物的 活化,激活后的阿科特则通过磷酸化作用激活MTOR等下游靶蛋白,从而促进 细胞的增殖、分化及迁移等
4 致病机制 标题文本预设 点击此处更换文本点击此处更换文本点 击此处更换文本 EGFR 作为表皮生长因子家族中的一员,能够激活许多经典的信号转导通 路,其中重要的一条就是 PI3K/Akt/mTOR 通路。 PI3K是一种可催化磷脂酰肌醇D3位磷酸化的脂类激酶,可以通过与具有磷 酸化酪氨酸残基的生长因子受体(如表皮生长因子受体、c-Met等)或连接 蛋白相互作用,导致二聚体构象的转变而导致PI3K的活化,活化后的PI3K将 导致PIP2转化为PIP3PIP3作为第二信使会进而激活各类下游蛋白[第一条 第二款)而阿科特即是PI3K下游的关键蛋白之一.阿科特家族的成员有三个, 即Aktl、Akt2和AKT3它们在结构和功能方面很相似,均参与了下游底物的 活化,激活后的阿科特则通过磷酸化作用激活MTOR等下游靶蛋白,从而促进 细胞的增殖、分化及迁移等