
(2)消泡油添加过量 (3)生理酸性物质的存在 4,引起 pH 上升的因素(1)氮源过多 (2)生理碱性物质的存在 (3)中间补料,碱性物质添加过多 5,最适 pH 的选择选择原则:有利于菌体生长和产物的合成。一般根据实 验结果确定。 最适 pH 与菌株,培养基组成,发酵工艺有关。应按发酵过程的不同阶段分别控 制不同的 pH 范围。 最适 pH 与微生物生长,产物形成之间相互关系有四种类型: (1)菌体比生长速率μ和产物比生产速率 QP 的最适 pH 在一个相似的较宽 的范围内(比较容易控制); (2)μ较宽, Qp 范围较窄,或μ较窄, Qp 范围较宽(难控制,应严格控 制); (3)μ和 Qp 对 pH 都很敏感,其最适 pH 相同(应严格控制); (4)更复杂,μ和 Qp 对 pH 都很敏感,并有各自的最适 pH(难度最大); 6,pH 的控制在发酵过程中可通过以下方式来控制 pH (1)调节基础培养基的配方 (2)调节碳氮比(C/N) (3)添加缓冲剂 (4)补料控制 (5)直接加酸加碱 (6)补加碳源或氮源 三、氧对发酵的影响 大多数发酵过程是好氧的,因此需要供氧。如果考虑呼吸的化学计量,则葡 萄糖的氧化可由下式表示: C6H 12O6 十 6O2=6H2O 十 6CO2 只有当这两种反应物均溶于水后,才对菌体有用。 氧在水中的溶解度比葡萄糖要小约 6000 倍左右(氧在水中的饱和度约为 l0mg/L) 。许多发酵的生产能力受到氧利用限制,因此氧成为影响发酵效率的重 要因素。1,发酵过程中氧的需求 尽管考虑了呼吸的化学计量而使供氧问题得以正确评价,但由于未曾将转化 为生物物质的碳加以考虑而使菌体的真实需氧情况难以表明。许多研究工作者已 经考虑到氧、碳源、氮源转化为生物物质的总化学计量关系,并利用这样的关系 来预测发酵的需氧情况。从这些测定结果发现菌体的需氧似乎完全取决于培养基 中的碳源。Darlington(1964)以 C3.92H6.5O1.94 来表述 100 克酵母菌体(干重)的组分, 并对由碳氢化合物和碳水化合物生产酵母推导得到如下方程式: 根据 Darlington 方程式,可知同样生 100g 菌体,用碳氢化合物所需的氧约 6.67CH2O + 2.1O2 = C3.92H6.5O1.94 + 2.75CO2 +3.42H2O 7.14CH2 + 6.135O2 = C3.92H6.5O1.94 +3.22CO2 +3.89H2O
(2)消泡油添加过量 (3)生理酸性物质的存在 4,引起 pH 上升的因素(1)氮源过多 (2)生理碱性物质的存在 (3)中间补料,碱性物质添加过多 5,最适 pH 的选择选择原则:有利于菌体生长和产物的合成。一般根据实 验结果确定。 最适 pH 与菌株,培养基组成,发酵工艺有关。应按发酵过程的不同阶段分别控 制不同的 pH 范围。 最适 pH 与微生物生长,产物形成之间相互关系有四种类型: (1)菌体比生长速率μ和产物比生产速率 QP 的最适 pH 在一个相似的较宽 的范围内(比较容易控制); (2)μ较宽, Qp 范围较窄,或μ较窄, Qp 范围较宽(难控制,应严格控 制); (3)μ和 Qp 对 pH 都很敏感,其最适 pH 相同(应严格控制); (4)更复杂,μ和 Qp 对 pH 都很敏感,并有各自的最适 pH(难度最大); 6,pH 的控制在发酵过程中可通过以下方式来控制 pH (1)调节基础培养基的配方 (2)调节碳氮比(C/N) (3)添加缓冲剂 (4)补料控制 (5)直接加酸加碱 (6)补加碳源或氮源 三、氧对发酵的影响 大多数发酵过程是好氧的,因此需要供氧。如果考虑呼吸的化学计量,则葡 萄糖的氧化可由下式表示: C6H 12O6 十 6O2=6H2O 十 6CO2 只有当这两种反应物均溶于水后,才对菌体有用。 氧在水中的溶解度比葡萄糖要小约 6000 倍左右(氧在水中的饱和度约为 l0mg/L) 。许多发酵的生产能力受到氧利用限制,因此氧成为影响发酵效率的重 要因素。1,发酵过程中氧的需求 尽管考虑了呼吸的化学计量而使供氧问题得以正确评价,但由于未曾将转化 为生物物质的碳加以考虑而使菌体的真实需氧情况难以表明。许多研究工作者已 经考虑到氧、碳源、氮源转化为生物物质的总化学计量关系,并利用这样的关系 来预测发酵的需氧情况。从这些测定结果发现菌体的需氧似乎完全取决于培养基 中的碳源。Darlington(1964)以 C3.92H6.5O1.94 来表述 100 克酵母菌体(干重)的组分, 并对由碳氢化合物和碳水化合物生产酵母推导得到如下方程式: 根据 Darlington 方程式,可知同样生 100g 菌体,用碳氢化合物所需的氧约 6.67CH2O + 2.1O2 = C3.92H6.5O1.94 + 2.75CO2 +3.42H2O 7.14CH2 + 6.135O2 = C3.92H6.5O1.94 +3.22CO2 +3.89H2O

为用碳水化合物生产时的 3 倍。 Johnson(1964)得出如下方程: A 为燃烧 1g 底物成 CO2、H2O 和 NH3(如底物中有氮存在)所需氧的量, 此值易于计算获得;B 为燃烧 1g 菌体成 CO2、H2O 和 NH3 所需氧的量,如果 菌体组分已知的话,则也可计算获得。 C 为生产 1 克菌体所需氧的量; Y 为 1 克底物转化成菌体的克数。 因此,A/Y 为燃烧生成 1g 菌体的底物所需的氧,而 B 为燃烧菌体所需氧的 量;它们之间的差为 C,即为转化底物成菌体所需氧的量。 将 Johnson 方程式 应用于利用葡萄糖和烷烃生产酵母的下列方程式为: 如果对葡萄糖来说 Y 值取 50%,而对烷烃来说 Y 值取 100%; 则:C 对葡萄糖 =24.95mmol 氧/g 菌体; C 对烷烃 =65.4mmol 氧/g 菌体 Mateles(1971)推导得一种碳源与需 氧间关系的方程式,他假定代谢产物仅为菌体、CO2、H2O;以及菌体的正常组 分 C 为 53%、N 为 12%、O 为 19%、H 为 7%,则 Mateles 利用此关系式对许多菌体利用各种基质所需的氧进行了计算 生长 于不同基质上的不同微生物的需氧要求 基 质 微生物 g 氧/g 干菌体 葡萄糖 甲 醇 辛 烷 大肠杆菌 假单孢菌 C 假单孢菌 0.4 1.2 1.7 上述方程式与生物物质的形成有关,并假定微生物除产生水、二氧化碳外, 无其它产物形成。因此,这些方程仅能适用于菌体生产过程中预测氧的要求,而 对菌体能形成其它产物的过程则需要加以修正。例如 Cooney(1979)建议用下列方 程式计算青霉素发酵: Y B C A − = / 41.7 / 41.7 101.7 33.33 = − = − Y Y C mmol g C mmol g 对烷烃 ( ) 对葡萄糖 ( ) :碳源的分子量 :与碳源有关的菌体生成量 菌体形成的克数 耗氧的克数 M Y Y M C H O 1.58 32 8 16 = − + − :形成的卞青霉素钠的克数 :形成的干菌体克数 :消耗每克葡萄糖所形成的卞青霉素钠的克数 形成每克卞青霉素钠所需氧的克数 P X Y Y Y Y P O O P P X O P P O / / 0.5 3 0.6 / : 0.43 / = − −
为用碳水化合物生产时的 3 倍。 Johnson(1964)得出如下方程: A 为燃烧 1g 底物成 CO2、H2O 和 NH3(如底物中有氮存在)所需氧的量, 此值易于计算获得;B 为燃烧 1g 菌体成 CO2、H2O 和 NH3 所需氧的量,如果 菌体组分已知的话,则也可计算获得。 C 为生产 1 克菌体所需氧的量; Y 为 1 克底物转化成菌体的克数。 因此,A/Y 为燃烧生成 1g 菌体的底物所需的氧,而 B 为燃烧菌体所需氧的 量;它们之间的差为 C,即为转化底物成菌体所需氧的量。 将 Johnson 方程式 应用于利用葡萄糖和烷烃生产酵母的下列方程式为: 如果对葡萄糖来说 Y 值取 50%,而对烷烃来说 Y 值取 100%; 则:C 对葡萄糖 =24.95mmol 氧/g 菌体; C 对烷烃 =65.4mmol 氧/g 菌体 Mateles(1971)推导得一种碳源与需 氧间关系的方程式,他假定代谢产物仅为菌体、CO2、H2O;以及菌体的正常组 分 C 为 53%、N 为 12%、O 为 19%、H 为 7%,则 Mateles 利用此关系式对许多菌体利用各种基质所需的氧进行了计算 生长 于不同基质上的不同微生物的需氧要求 基 质 微生物 g 氧/g 干菌体 葡萄糖 甲 醇 辛 烷 大肠杆菌 假单孢菌 C 假单孢菌 0.4 1.2 1.7 上述方程式与生物物质的形成有关,并假定微生物除产生水、二氧化碳外, 无其它产物形成。因此,这些方程仅能适用于菌体生产过程中预测氧的要求,而 对菌体能形成其它产物的过程则需要加以修正。例如 Cooney(1979)建议用下列方 程式计算青霉素发酵: Y B C A − = / 41.7 / 41.7 101.7 33.33 = − = − Y Y C mmol g C mmol g 对烷烃 ( ) 对葡萄糖 ( ) :碳源的分子量 :与碳源有关的菌体生成量 菌体形成的克数 耗氧的克数 M Y Y M C H O 1.58 32 8 16 = − + − :形成的卞青霉素钠的克数 :形成的干菌体克数 :消耗每克葡萄糖所形成的卞青霉素钠的克数 形成每克卞青霉素钠所需氧的克数 P X Y Y Y Y P O O P P X O P P O / / 0.5 3 0.6 / : 0.43 / = − −

但是,由于菌体的代谢是受发酵液中的溶氧浓度的影响,故简单地依据总需 求来供氧是欠妥的。溶氧浓度对比摄氧率(QO2 每克干菌体每小时所消耗的氧的 毫摩尔数)用米氏型曲线表示。 临界氧浓度(C 临):指不影响菌体呼吸所允许的最低氧浓度,或微生物对发 酵液中溶解氧浓度的最低要求。某些微生物的临界氧浓度 微 生 物 温度(˚C) 临界氧浓度(mmol/L) 固 氮 菌 大肠杆菌 酵 母 产黄青霉 30 37 30 24 0.018 0.008 0.004 0.022 2、溶氧对发酵的影响由此可知,只有使溶氧浓度高于其临界值,才能维持 菌体的最大比摄氧率,得到最大的菌体合成量。如果溶氧浓度低于临界值,则菌 体代谢受到干扰。但是,发酵工业的目标是要得到菌体发酵的产物而不是菌体本 身。因此,由氧饥饿而引起的细胞代谢干扰,可能对形成某些产物是有利的;同 理,当提供的氧浓度远大于临界值时,虽对菌体形成无妨,但也许能刺激产物的 形成;所以,某种产物形成的最佳条件可能不同于菌体生长的最佳通气条件。 根 据需氧不同,可将初级代谢发酵分为: a.供氧充足条件下,产量最大;若供氧不足,合成受强烈抑制; 如:谷氨酸,精氨酸,脯氨酸等 b.供氧充足条件下,可得最高产量;若供氧受限,产量受影响不明显; 如:异亮氨酸,赖氨酸,苏氨酸等 c.若供氧受限,细胞呼吸受抑制时,才获得最大量产物;若供氧充足,产物 形成反而受抑制; 如:亮氨酸,缬氨酸,苯丙氨酸等但在实际生产过程中需注意: –溶解氧浓度过低(代谢异常,产量降低) Dissolved Oxygen Concentration Ccritical QO2
但是,由于菌体的代谢是受发酵液中的溶氧浓度的影响,故简单地依据总需 求来供氧是欠妥的。溶氧浓度对比摄氧率(QO2 每克干菌体每小时所消耗的氧的 毫摩尔数)用米氏型曲线表示。 临界氧浓度(C 临):指不影响菌体呼吸所允许的最低氧浓度,或微生物对发 酵液中溶解氧浓度的最低要求。某些微生物的临界氧浓度 微 生 物 温度(˚C) 临界氧浓度(mmol/L) 固 氮 菌 大肠杆菌 酵 母 产黄青霉 30 37 30 24 0.018 0.008 0.004 0.022 2、溶氧对发酵的影响由此可知,只有使溶氧浓度高于其临界值,才能维持 菌体的最大比摄氧率,得到最大的菌体合成量。如果溶氧浓度低于临界值,则菌 体代谢受到干扰。但是,发酵工业的目标是要得到菌体发酵的产物而不是菌体本 身。因此,由氧饥饿而引起的细胞代谢干扰,可能对形成某些产物是有利的;同 理,当提供的氧浓度远大于临界值时,虽对菌体形成无妨,但也许能刺激产物的 形成;所以,某种产物形成的最佳条件可能不同于菌体生长的最佳通气条件。 根 据需氧不同,可将初级代谢发酵分为: a.供氧充足条件下,产量最大;若供氧不足,合成受强烈抑制; 如:谷氨酸,精氨酸,脯氨酸等 b.供氧充足条件下,可得最高产量;若供氧受限,产量受影响不明显; 如:异亮氨酸,赖氨酸,苏氨酸等 c.若供氧受限,细胞呼吸受抑制时,才获得最大量产物;若供氧充足,产物 形成反而受抑制; 如:亮氨酸,缬氨酸,苯丙氨酸等但在实际生产过程中需注意: –溶解氧浓度过低(代谢异常,产量降低) Dissolved Oxygen Concentration Ccritical QO2
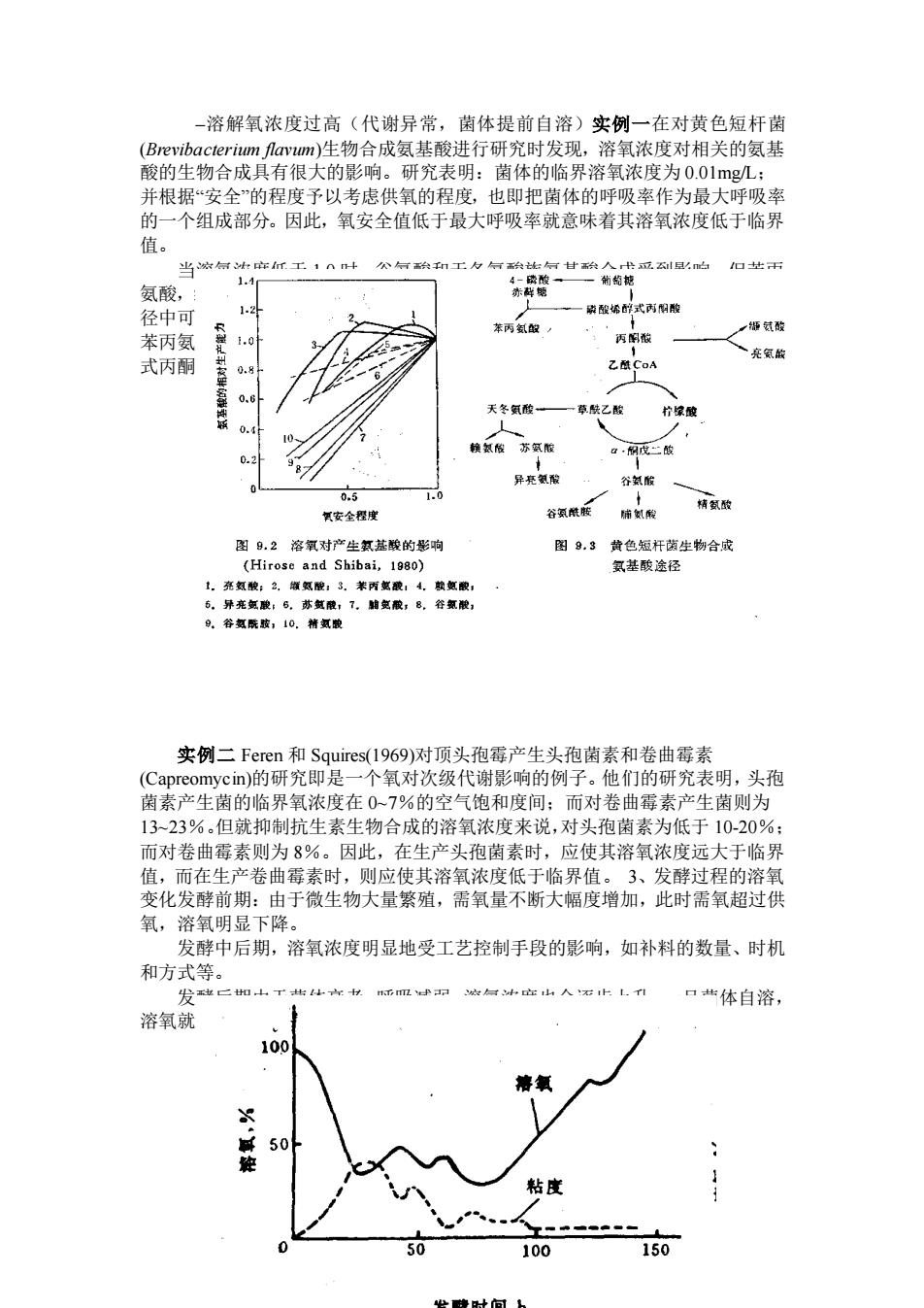
–溶解氧浓度过高(代谢异常,菌体提前自溶)实例一在对黄色短杆菌 (Brevibacterium flavum)生物合成氨基酸进行研究时发现,溶氧浓度对相关的氨基 酸的生物合成具有很大的影响。研究表明:菌体的临界溶氧浓度为 0.01mg/L; 并根据“安全”的程度予以考虑供氧的程度,也即把菌体的呼吸率作为最大呼吸率 的一个组成部分。因此,氧安全值低于最大呼吸率就意味着其溶氧浓度低于临界 值。 当溶氧浓度低于 1.0 时,谷氨酸和天冬氨酸族氨基酸合成受到影响,但苯丙 氨酸,缬氨酸和亮氨酸最佳合成的溶氧浓度分别为 0.55、0.60 和 0.85。从合成途 径中可知,谷氨酸和天冬氨酸族的氨基酸来自于三羧酸循环(TCA)的中间体,而 苯丙氨酸、缬氨酸和亮氨酸来自于糖酵解的中间体,即来自于丙酮酸和磷酸稀醇 式丙酮酸。 实例二 Feren 和 Squires(1969)对顶头孢霉产生头孢菌素和卷曲霉素 (Capreomycin)的研究即是一个氧对次级代谢影响的例子。他们的研究表明,头孢 菌素产生菌的临界氧浓度在 0~7%的空气饱和度间;而对卷曲霉素产生菌则为 13~23%。但就抑制抗生素生物合成的溶氧浓度来说,对头孢菌素为低于 10-20%; 而对卷曲霉素则为 8%。因此,在生产头孢菌素时,应使其溶氧浓度远大于临界 值,而在生产卷曲霉素时,则应使其溶氧浓度低于临界值。 3、发酵过程的溶氧 变化发酵前期:由于微生物大量繁殖,需氧量不断大幅度增加,此时需氧超过供 氧,溶氧明显下降。 发酵中后期,溶氧浓度明显地受工艺控制手段的影响,如补料的数量、时机 和方式等。 发酵后期由于菌体衰老,呼吸减弱,溶氧浓度也会逐步上升,一旦菌体自溶, 溶氧就会明显地上升
–溶解氧浓度过高(代谢异常,菌体提前自溶)实例一在对黄色短杆菌 (Brevibacterium flavum)生物合成氨基酸进行研究时发现,溶氧浓度对相关的氨基 酸的生物合成具有很大的影响。研究表明:菌体的临界溶氧浓度为 0.01mg/L; 并根据“安全”的程度予以考虑供氧的程度,也即把菌体的呼吸率作为最大呼吸率 的一个组成部分。因此,氧安全值低于最大呼吸率就意味着其溶氧浓度低于临界 值。 当溶氧浓度低于 1.0 时,谷氨酸和天冬氨酸族氨基酸合成受到影响,但苯丙 氨酸,缬氨酸和亮氨酸最佳合成的溶氧浓度分别为 0.55、0.60 和 0.85。从合成途 径中可知,谷氨酸和天冬氨酸族的氨基酸来自于三羧酸循环(TCA)的中间体,而 苯丙氨酸、缬氨酸和亮氨酸来自于糖酵解的中间体,即来自于丙酮酸和磷酸稀醇 式丙酮酸。 实例二 Feren 和 Squires(1969)对顶头孢霉产生头孢菌素和卷曲霉素 (Capreomycin)的研究即是一个氧对次级代谢影响的例子。他们的研究表明,头孢 菌素产生菌的临界氧浓度在 0~7%的空气饱和度间;而对卷曲霉素产生菌则为 13~23%。但就抑制抗生素生物合成的溶氧浓度来说,对头孢菌素为低于 10-20%; 而对卷曲霉素则为 8%。因此,在生产头孢菌素时,应使其溶氧浓度远大于临界 值,而在生产卷曲霉素时,则应使其溶氧浓度低于临界值。 3、发酵过程的溶氧 变化发酵前期:由于微生物大量繁殖,需氧量不断大幅度增加,此时需氧超过供 氧,溶氧明显下降。 发酵中后期,溶氧浓度明显地受工艺控制手段的影响,如补料的数量、时机 和方式等。 发酵后期由于菌体衰老,呼吸减弱,溶氧浓度也会逐步上升,一旦菌体自溶, 溶氧就会明显地上升

4、溶氧的控制 (1)调节通风与搅拌(具体内容在通风发酵设备中介绍) (2)限制基础培养基的浓度,使发酵器内的生物体浓度维持于适当水平; 并以补料方式供给某些营养成分而控制菌体生长率和呼吸率。 四、CO2对发酵的影响及控制 CO2 是微生物的代谢产物,同时也是某些合成代谢的基质。它是细胞代谢的 重要指标。在发酵过程中,CO2 有可能对发酵有促进作用,也有可能有抑制作用。 1、CO2 对发酵的影响 (1)CO2 对菌体具有抑制作用 当排气中 CO2 的浓度高于 4%时,微生物的糖代谢和呼吸速率下降。例如, 发酵液中 CO2 的浓度达到 1.6×10-1mol,就会严重抑制酵母的生长;当进气口 CO2 的含量占混合气体的 80%时,酵母活力与对照相比降低 20%。(2)CO2 对发 酵的影响 ①对发酵促进。如牛链球菌发酵生产多糖,最重要的发酵条件是提供的空气 中要含 5%的 CO2。 ②对发酵抑制。如对肌苷、异亮氨酸、组氨酸、抗生素等发酵的抑制 ③影响发酵液的酸碱平衡 2、CO2 对发酵影响的机理 CO2 及 HCO3 -主要是影响细胞膜的结构,导致膜的流动性及表面电荷密度发 生改变,影响到细胞膜的输送效率,导致细胞生长受到抑制、形态发生改变。 培养液中的CO2主要作用于细胞膜的脂质核心部位;HCO3 -影响细胞膜的膜蛋白。 3、CO2 的控制 CO2 在发酵液中的浓度变化不像溶解氧那样有一定的规律。它的大小受到许 多因素的影响,如细胞的呼吸强度、发酵液的流变学特性、通气搅拌程度、罐压 大小、设备规模等。在发酵过程中通常通过调节通风和搅拌来控制。 五、发酵过程中的泡沫及其控制 1,泡沫的性质 泡沫是气体被分散在少量液体中的胶体体系。泡沫间被一层液膜隔开而彼此 不相连通。发酵过程中所遇到的泡沫,其分散相是无菌空气和代谢气体,连续相 是发酵液。 2,泡沫的类型一类存在于发酵液的液面上,这类泡沫气相所占比例特别大, 并且泡沫与它下面的液体之间有能分辫的界线。如在某些稀薄的前期发酵液或种 子培养液中所见到的。 另一种泡沫是出现在粘稠的菌丝发酵液当中。这种泡沫分散很细,而且很均 匀,也较稳定。泡沫与液体间没有明显的波面界限,在鼓泡的发酵液中气体分散
4、溶氧的控制 (1)调节通风与搅拌(具体内容在通风发酵设备中介绍) (2)限制基础培养基的浓度,使发酵器内的生物体浓度维持于适当水平; 并以补料方式供给某些营养成分而控制菌体生长率和呼吸率。 四、CO2对发酵的影响及控制 CO2 是微生物的代谢产物,同时也是某些合成代谢的基质。它是细胞代谢的 重要指标。在发酵过程中,CO2 有可能对发酵有促进作用,也有可能有抑制作用。 1、CO2 对发酵的影响 (1)CO2 对菌体具有抑制作用 当排气中 CO2 的浓度高于 4%时,微生物的糖代谢和呼吸速率下降。例如, 发酵液中 CO2 的浓度达到 1.6×10-1mol,就会严重抑制酵母的生长;当进气口 CO2 的含量占混合气体的 80%时,酵母活力与对照相比降低 20%。(2)CO2 对发 酵的影响 ①对发酵促进。如牛链球菌发酵生产多糖,最重要的发酵条件是提供的空气 中要含 5%的 CO2。 ②对发酵抑制。如对肌苷、异亮氨酸、组氨酸、抗生素等发酵的抑制 ③影响发酵液的酸碱平衡 2、CO2 对发酵影响的机理 CO2 及 HCO3 -主要是影响细胞膜的结构,导致膜的流动性及表面电荷密度发 生改变,影响到细胞膜的输送效率,导致细胞生长受到抑制、形态发生改变。 培养液中的CO2主要作用于细胞膜的脂质核心部位;HCO3 -影响细胞膜的膜蛋白。 3、CO2 的控制 CO2 在发酵液中的浓度变化不像溶解氧那样有一定的规律。它的大小受到许 多因素的影响,如细胞的呼吸强度、发酵液的流变学特性、通气搅拌程度、罐压 大小、设备规模等。在发酵过程中通常通过调节通风和搅拌来控制。 五、发酵过程中的泡沫及其控制 1,泡沫的性质 泡沫是气体被分散在少量液体中的胶体体系。泡沫间被一层液膜隔开而彼此 不相连通。发酵过程中所遇到的泡沫,其分散相是无菌空气和代谢气体,连续相 是发酵液。 2,泡沫的类型一类存在于发酵液的液面上,这类泡沫气相所占比例特别大, 并且泡沫与它下面的液体之间有能分辫的界线。如在某些稀薄的前期发酵液或种 子培养液中所见到的。 另一种泡沫是出现在粘稠的菌丝发酵液当中。这种泡沫分散很细,而且很均 匀,也较稳定。泡沫与液体间没有明显的波面界限,在鼓泡的发酵液中气体分散