
一、结构与装配 。 根据等电点分3类:a-actin分布于肌细胞;B-和y-分 布于所有细胞。 ·单体呈哑铃形,称G-actin;多聚体称F-actin。 结构高度保守,酵母和兔子actin有88%的同源性。 ·需要翻译后修饰,如N-端乙酰化或组氨酸残基的甲 基化。 Dr Tian 2008
一、结构与装配 • 根据等电点分 3类:α‐actin分布于肌细胞;β‐ 和γ‐ 分 布于所有细胞。 • 单体呈哑铃形,称 G ‐actin;多聚体称 F ‐actin 。 • 结构高度保守,酵母和兔子actin 有88%的同源性。 • 需要翻译后修饰,如 N ‐端乙酰化或组氨酸残基的甲 基化

微丝的装配 ·条件:ATP、适宜温度、K+和Mg2+。 ·过程:2-3个actin聚集成核心(核化);ATP-actin 分子向核心两端加合。 ·极性:ATP-actin加到(+)极的速度要比加到(-)极的 速度快5-10倍。 ·踏车行为:单体可同时在(+)端添加,在()端分离
微丝的装配 • 条件:ATP、适宜温度、K+和Mg2+。 • 过程:2‐3个actin聚集成核心(核化);ATP‐actin 分子向核心两端加合。 • 极性:ATP‐actin加到(+)极的速度要比加到(‐)极的 速度快5‐10倍。 • 踏车行为:单体可同时在(+)端添加,在(‐)端分离
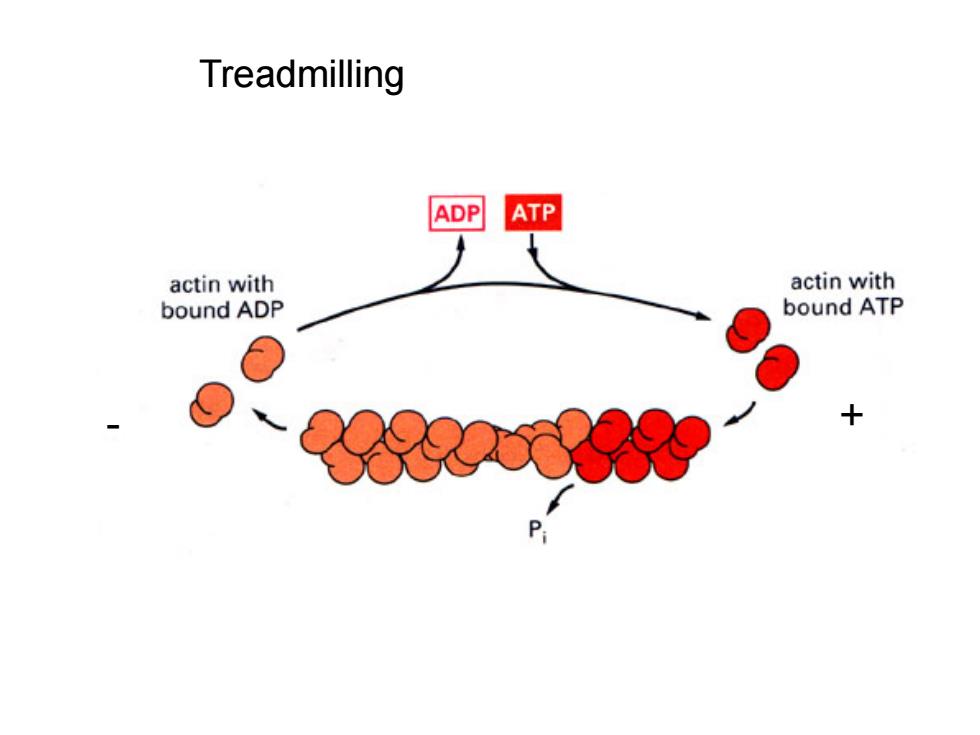
Treadmilling ADP ATP actin with actin with bound ADP bound ATP
Treadmilling - +

·细胞中大多数微丝结构处于动态组装和去组装过 程中,并通过这种方式实现其功能。 ●至 细胞松弛素(cytochalasin)可切断微丝纤维,并 结合在微丝末端抑制肌动蛋白加合到微丝纤维上。 鬼笔环肽(phalloidin)与微丝能够特异性的结合, 使微丝纤维稳定而抑制其功能。荧光标记的鬼笔 环肽可特异性的显示微丝。 D7am2008
• 细胞中大多数微丝结构处于动态组装和去组装过 程中,并通过这种方式实现其功能。 • 细胞松弛素(cytochalasin)可切断微丝纤维,并 结合在微丝末端抑制肌动蛋白加合到微丝纤维上。 • 鬼笔环肽(phalloidin)与微丝能够特异性的结合, 使微丝纤维稳定而抑制其功能。荧光标记的鬼笔 环肽可特异性的显示微丝

二、微丝结合蛋白 已知的的微丝结合蛋白有100多种, 1.核化蛋白:使游离actin核化,开始组装,Arp ·2.单体隐蔽蛋白:阻止游离actin向纤维添加,thymosin 3. 封端蛋白:使纤维稳定,capZ ·4.单体聚合蛋白:将结合的单体安装到纤维,profilin 5.微丝解聚蛋白:cofilin 6.交联蛋白:fimbrin 。7.纤维切断蛋白:gelsolin ·8.膜结合蛋白:vinculin Dr Tian 2008
二、微丝结合蛋白 • 已知的的微丝结合蛋白有100多种, • 1 .核化蛋白:使游离actin核化,开始组装,Arp • 2.单体隐蔽蛋白:阻止游离actin向纤维添加,thymosin • 3.封端蛋白:使纤维稳定,Cap Z • 4.单体聚合蛋白:将结合的单体安装到纤维,profilin • 5.微丝解聚蛋白:cofilin • 6.交联蛋白:fimbrin • 7.纤维切断蛋白:gelsolin • 8.膜结合蛋白:vinculin