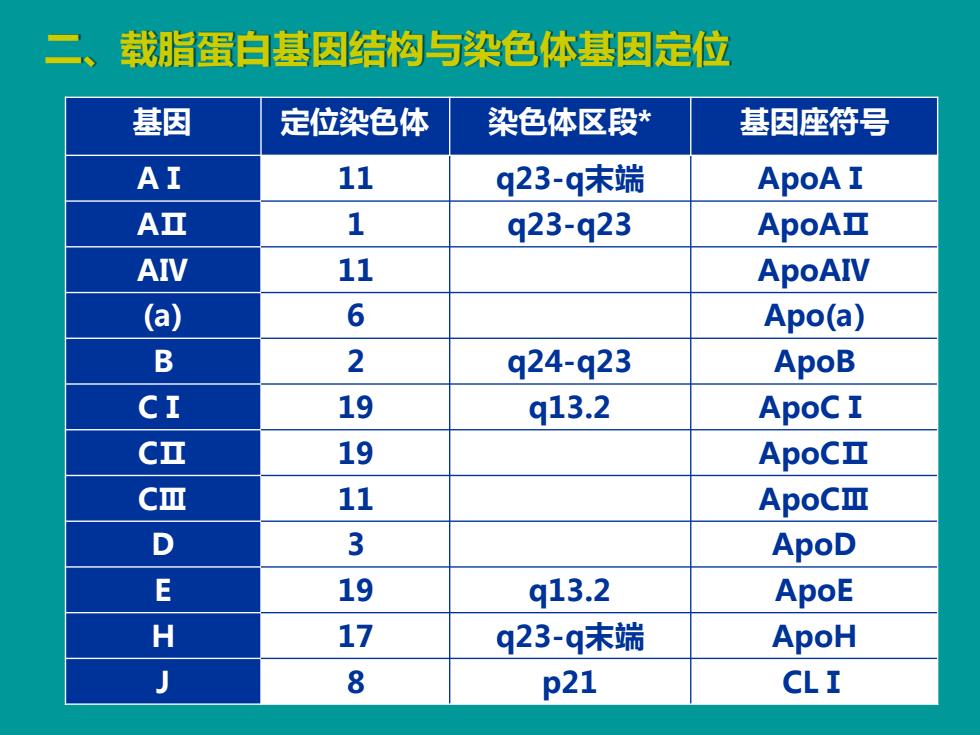
载脂蛋白基因结构与染色体基因定位 基因 定位染色体 染色体区段* 基因座符号 AI 11 q23-q末端 ApoA I AⅡ 1 q23-q23 ApoAII AIV 11 ApoAIV (a) 6 Apo(a) B 2 q24-q23 ApoB CI 19 q13.2 ApoC I C亚 19 ApoCI CⅢ 11 ApoCⅢ D 3 ApoD E 19 q13.2 ApoE H 17 q23-q末端 ApoH 8 p21 CLI
基因 定位染色体 染色体区段* 基因座符号 AⅠ 11 q23-q末端 ApoAⅠ AⅡ 1 q23-q23 ApoAⅡ AⅣ 11 ApoAⅣ (a) 6 Apo(a) B 2 q24-q23 ApoB CⅠ 19 q13.2 ApoCⅠ CⅡ 19 ApoCⅡ CⅢ 11 ApoCⅢ D 3 ApoD E 19 q13.2 ApoE H 17 q23-q末端 ApoH J 8 p21 CLⅠ 二、载脂蛋白基因结构与染色体基因定位

第三节脂蛋白受体 脂类在血液中以脂蛋白形式进行运送 并可与细胞膜上存在的特异受体相结合, 被摄取进入细胞内进行代谢。 迄今为止报道的受体已有很多种,主要 有LDL受体、清道夫受体、VLDL受体。 一、低密度脂蛋白受体 二、极低密度脂蛋白受体 三、清道夫受体
脂类在血液中以脂蛋白形式进行运送, 并可与细胞膜上存在的特异受体相结合, 被摄取进入细胞内进行代谢。 迄今为止报道的受体已有很多种,主要 有LDL受体、清道夫受体、VLDL受体。 第三节 脂蛋白受体 一、低密度脂蛋白受体 二、极低密度脂蛋白受体 三、清道夫受体

一、低密度脂蛋白受体 LDL受体结构 LDL受体(LDLR)是一种多功能蛋白,由836个 氨基酸残基组成36面体结构蛋白,分子量约 115kU,LDLR由五种不同结构域组成LDL受体对 含ApoB1O0的LDL,含ApoE的VLDL、B-VLDL、 VLDL残粒均有高亲和性
LDL受体(LDLR)是一种多功能蛋白,由836个 氨基 酸 残基组成36面体结构蛋白, 分子量约 115kU, LDLR由五种不同结构域组成 LDL受体对 含ApoB100 的LDL,含ApoE 的VLDL、 β-VLDL、 VLDL残粒均有高亲和性。 一、低密度脂蛋白受体 LDL受体结构
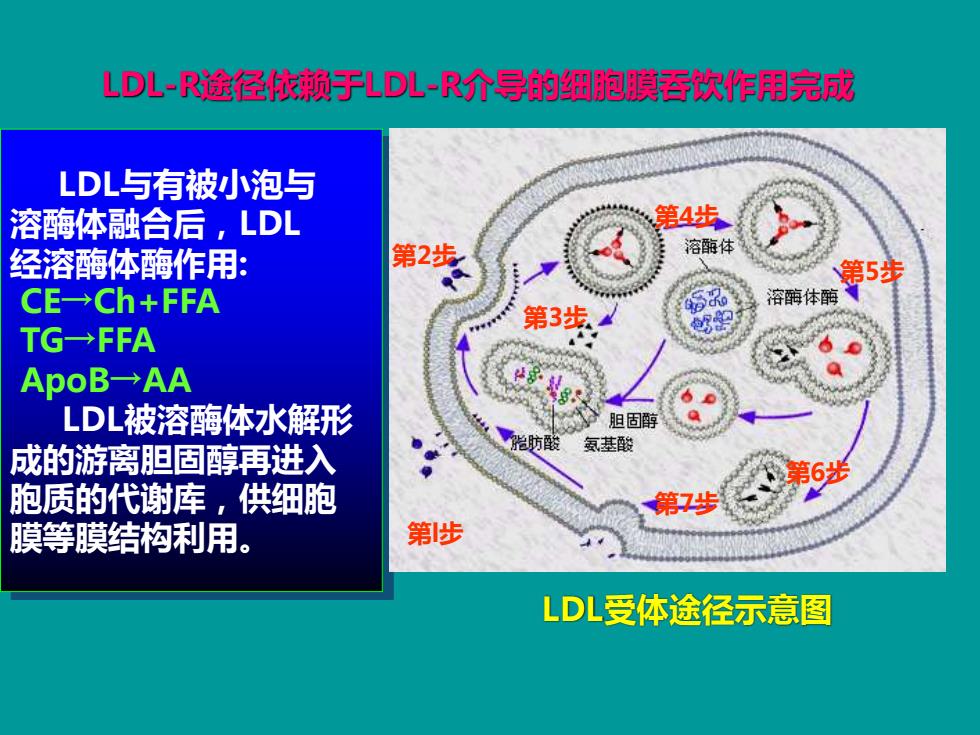
LD儿-R途径依赖于LD儿~R介导的细胞膜吞饮作用完成 LDL与有被小泡与 溶酶体融合后,LDL 第4步 经溶酶体酶作用: 第2步 溶酶体 第5步 CE→Ch+FFA 溶酶体酶、 第3步4 TG→FFA ApoB→AA LDL被溶酶体水解形 胆固醇 氨基酸 成的游离胆固醇再进入 胞质的代谢库,供细胞 第7步 膜等膜结构利用。 第步 LDL受体途径示意图
第l步 第2步 第3步 第4步 第5步 第6步 第7步 LDL受体途径示意图 LDL-R途径依赖于LDL-R介导的细胞膜吞饮作用完成 LDL与有被小泡与 溶酶体融合后,LDL 经溶酶体酶作用: CE→Ch+FFA TG→FFA ApoB→AA LDL被溶酶体水解形 成的游离胆固醇再进入 胞质的代谢库,供细胞 膜等膜结构利用

细胞内胆固醇代谢调节的重要作用: 若胞内浓度升高,可能出现抑制HMGCoA 还原酶,以减少自身的胆固醇合成,抑制 LDLR基因表达,减少LDLR的合成,从而减 少LDL的摄取,这种LDLR减少的调节过程称 为下调。 激活内质网ACAT,Ch一→CE,供细胞的需 要,经上述的变化,用以控制细胞内胆固醇 含量处于正常动态平衡状态.。 LDL65%~70%是依赖肝细胞的LDLR清 除
细胞内胆固醇代谢调节的重要作用: 若胞内浓度升高,可能出现抑制HMGCoA 还原酶,以减少自身的胆固醇合成,抑制 LDLR基因表达,减少LDLR的合成,从而减 少LDL的摄取,这种LDLR减少的调节过程称 为下调。 激活内质网ACAT, Ch→CE,供细胞的需 要,经上述的变化,用以控制细胞内胆固醇 含量处于正常动态平衡状态。 LDL65%~70%是依赖肝细胞的LDLR清 除