
在电子传递过程中,有几点需要说明 四种类型电子载体:黄素蛋白、细胞色素(含血红素辅 基)、Fe-S中心、辅酶Q。前三种与蛋白质结合,辅酶Q为 脂溶性醌。 电子传递起始于NADH脱氢酶催化NADH氧化,形成高能 电子(能量转化), 终止于O2形成水。 电 子 传 递 方 向 按 氧 化 还 原 电 势 递 增 的 方 向 传 递 (NAD+ /NAD最低,H2O/O2最高) 高能电子释放的能量驱动线粒体内膜三大复合物(H+ -泵) 将H+从基质侧泵到膜间隙, 形成跨线粒体内膜H+梯度(能 量转化) 电子传递链各组分在膜上不对称分布
在电子传递过程中,有几点需要说明 四种类型电子载体:黄素蛋白、细胞色素(含血红素辅 基)、Fe-S中心、辅酶Q。前三种与蛋白质结合,辅酶Q为 脂溶性醌。 电子传递起始于NADH脱氢酶催化NADH氧化,形成高能 电子(能量转化), 终止于O2形成水。 电 子 传 递 方 向 按 氧 化 还 原 电 势 递 增 的 方 向 传 递 (NAD+ /NAD最低,H2O/O2最高) 高能电子释放的能量驱动线粒体内膜三大复合物(H+ -泵) 将H+从基质侧泵到膜间隙, 形成跨线粒体内膜H+梯度(能 量转化) 电子传递链各组分在膜上不对称分布

ATP合成酶(磷酸化的分子基础) ATP合成酶的分布 分子结构 线粒体ATP合成系统的解离与重建实验证明电子传递与ATP 合成是由两个不 同的结构体系执行, F1颗粒具有ATP酶活性 工作特点:可逆性复合酶,即既能利用质子电化学梯度储存 的能量合成ATP, 又能水解ATP将质子从基质泵到膜间隙 ATP合成机制—Banding Change Mechanism (Boyer 1979) 亚单位相对于亚单位旋转的直接实验证据
ATP合成酶(磷酸化的分子基础) ATP合成酶的分布 分子结构 线粒体ATP合成系统的解离与重建实验证明电子传递与ATP 合成是由两个不 同的结构体系执行, F1颗粒具有ATP酶活性 工作特点:可逆性复合酶,即既能利用质子电化学梯度储存 的能量合成ATP, 又能水解ATP将质子从基质泵到膜间隙 ATP合成机制—Banding Change Mechanism (Boyer 1979) 亚单位相对于亚单位旋转的直接实验证据
Bacterium H ATP ADP+P; Plasma- NADH O2 H2O NAD membrane Mitochondrion Outer membrane Intermembrane space + Matrix ATP ADP +Pi NADH O2 H2O NAD Inner membrane Chloroplast Outer Light Intermembrane membrane space H2O 02 NADPH Stroma ATP ADP Inner membrane Thylakoid membrane

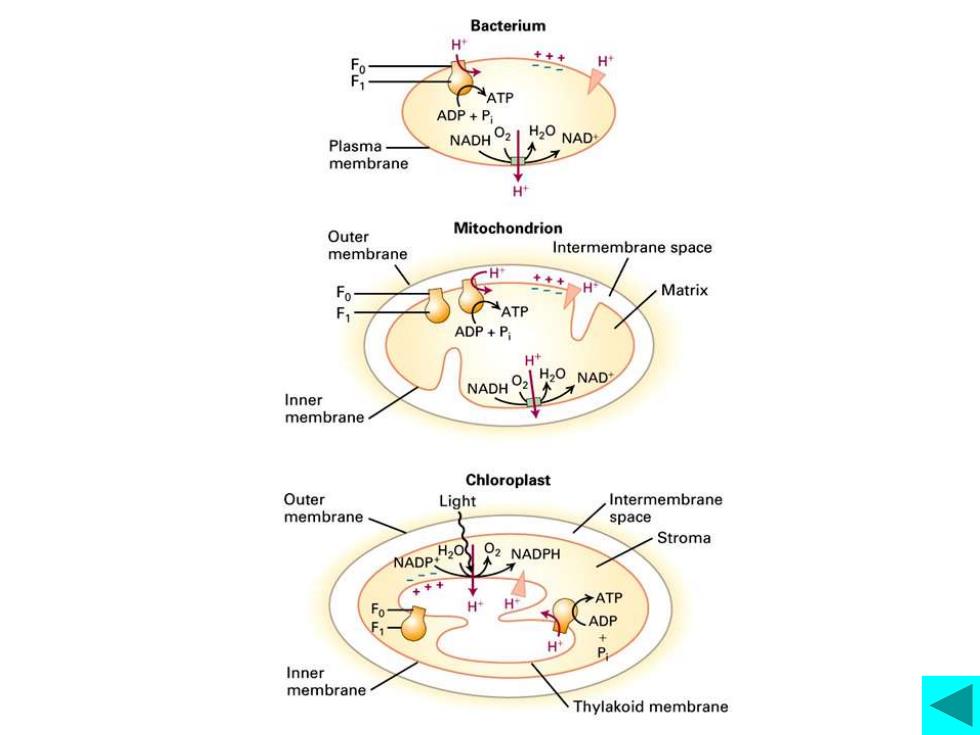
氧化磷酸化的偶联机制—化学渗透假说 化学渗透假说内容: 电子传递链各组分在线粒体内膜中不对称分布,当高能 电子沿其传递时,所释放的能量将H+从基质泵到膜间隙, 形成H+电化学梯度。在这个梯度驱使下,H+穿过ATP合成酶 回到基质,同时合成ATP,电化学梯度中蕴藏的能量储存到 ATP高能磷酸键。 质子动力势(proton motive force) 支持化学渗透假说的实验证据该实验表明: ·质子动力势乃ATP合成的动力 ·膜应具有完整性 ·电子传递与ATP合成是两件相关而又不同的事件
氧化磷酸化的偶联机制—化学渗透假说 化学渗透假说内容: 电子传递链各组分在线粒体内膜中不对称分布,当高能 电子沿其传递时,所释放的能量将H+从基质泵到膜间隙, 形成H+电化学梯度。在这个梯度驱使下,H+穿过ATP合成酶 回到基质,同时合成ATP,电化学梯度中蕴藏的能量储存到 ATP高能磷酸键。 质子动力势(proton motive force) 支持化学渗透假说的实验证据该实验表明: ·质子动力势乃ATP合成的动力 ·膜应具有完整性 ·电子传递与ATP合成是两件相关而又不同的事件

质子动力势的其他作用 物质转运 产热:冬眠动物与新生儿的Brown Fat Cell 线粒体产生大量热量
质子动力势的其他作用 物质转运 产热:冬眠动物与新生儿的Brown Fat Cell 线粒体产生大量热量