
1972 金花苔(原始不育株) 48A X 金花苔(何育株) 48B ⑧ 48A(不育株率62.5%) ×金花苔48B 1973 F (选不育株率高的株系回交) 、⑧ 48A(不育株率71.7%) BC 金花苔48B 1974 (选全不育的株系回交,其余淘汰) 、图 48A(不育株率88.9%) BC2 金花苔48B 1975 (选全不育的株系回交,其余淘汰) ⑧ 48A(不育株率94.5%) BC3 金花苔48B 1976 (选全不育的株系回交,其余海汰) ⑧ 48A(不育株率稳定在95%以上) 金花苔48B×金花苔48B 1977 (选全不育的株系回交,其余淘汰) 不育系 保特系 4) 金花苔不育系和保持系选育过程 Chapter 5 Self-incompatibility Self-incompatibility denotes the inability of a plant to set seeds upon self-pollination, despite the fact that both ovule and pollen development are normal and viable.This is due to some physiological hindrance to fertilization,for example slow pollen tube growth.Self-incompatibility differs from male sterility. The important differences are given in Table 5.1 Table 5.1 Differences between male sterility and self-incompatibility Male sterility self-incompatibility 1.Due to failure of processes such as pollen Due to some physiological hindrance to development fertilization.Failure to obtain seeds is due to slow pollen tube growth. 2.Pollen are non-functional(abortive) Pollen and ovule are normal and functional. 3.Genetic,cytoplasmic or both cytoplasmic Genetic and physiological factors and genetic factors control the expression of control the expression of male sterility self-incompatibility 1.Systems of Self-incompatibility Factors such as genetics,physiology and floral morphology determine the system of self-incompatibility. 16
16 4) 金花苔不育系和保持系选育过程 Chapter 5 Self-incompatibility Self-incompatibility denotes the inability of a plant to set seeds upon self-pollination, despite the fact that both ovule and pollen development are normal and viable. This is due to some physiological hindrance to fertilization, for example slow pollen tube growth.Self-incompatibility differs from male sterility. The important differences are given in Table 5.1 Table 5.1 Differences between male sterility and self-incompatibility Male sterility self-incompatibility 1.Due to failure of processes such as pollen development 2.Pollen are non-functional(abortive) 3.Genetic,cytoplasmic or both cytoplasmic and genetic factors control the expression of male sterility Due to some physiological hindrance to fertilization. Failure to obtain seeds is due to slow pollen tube growth. Pollen and ovule are normal and functional. Genetic and physiological factors control the expression of self-incompatibility 1.Systems of Self-incompatibility Factors such as genetics, physiology and floral morphology determine the system of self-incompatibility
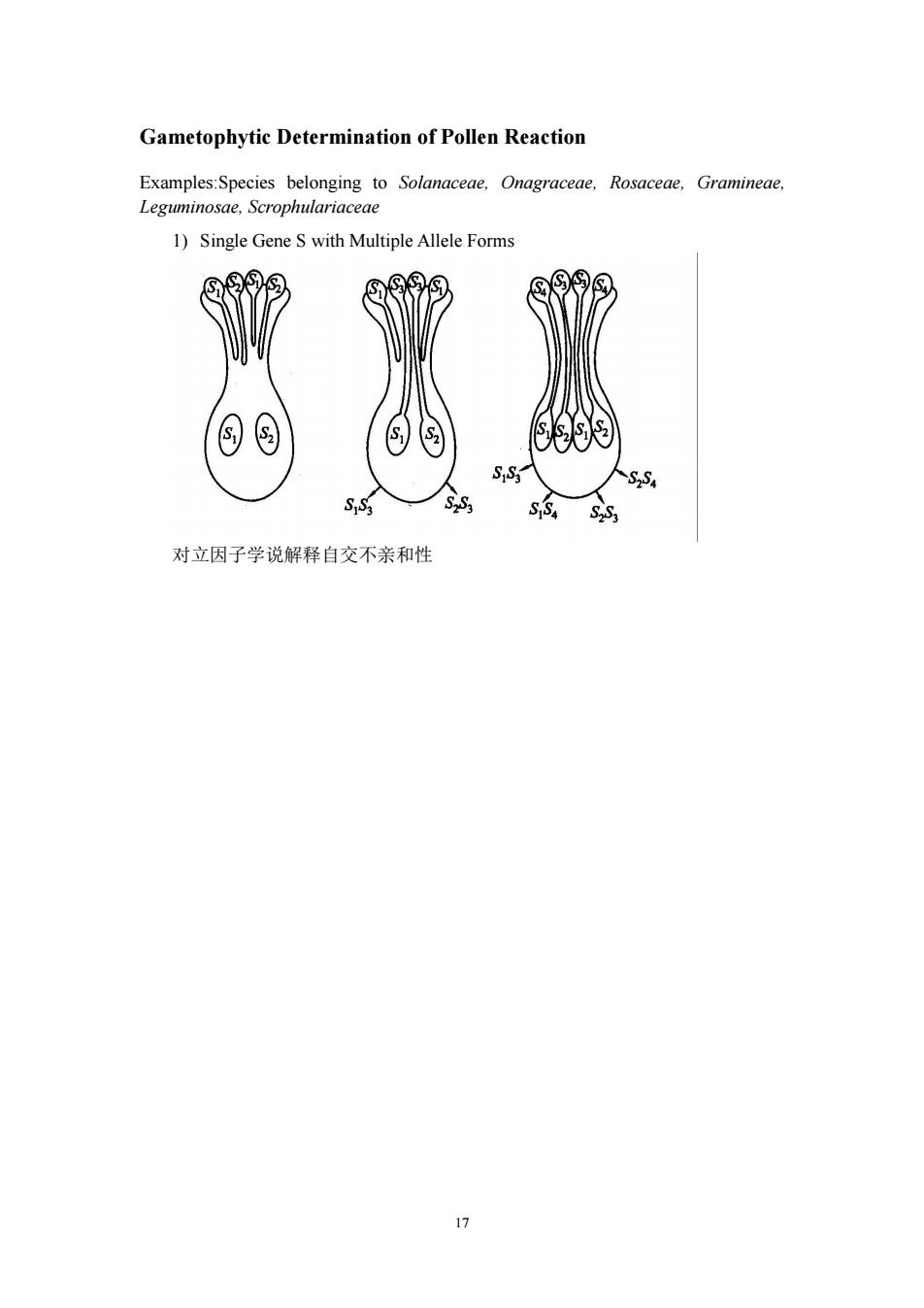
Gametophytic Determination of Pollen Reaction Examples:Species belonging to Solanaceae,Onagraceae, Rosaceae,Gramineae, Leguminosae,Scrophulariaceae 1)Single Gene S with Multiple Allele Forms ⑨ 8 S S S2S4 S:S3 对立因子学说解释自交不亲和性 公
17 Gametophytic Determination of Pollen Reaction Examples:Species belonging to Solanaceae, Onagraceae, Rosaceae, Gramineae, Leguminosae, Scrophulariaceae 1) Single Gene S with Multiple Allele Forms 对立因子学说解释自交不亲和性
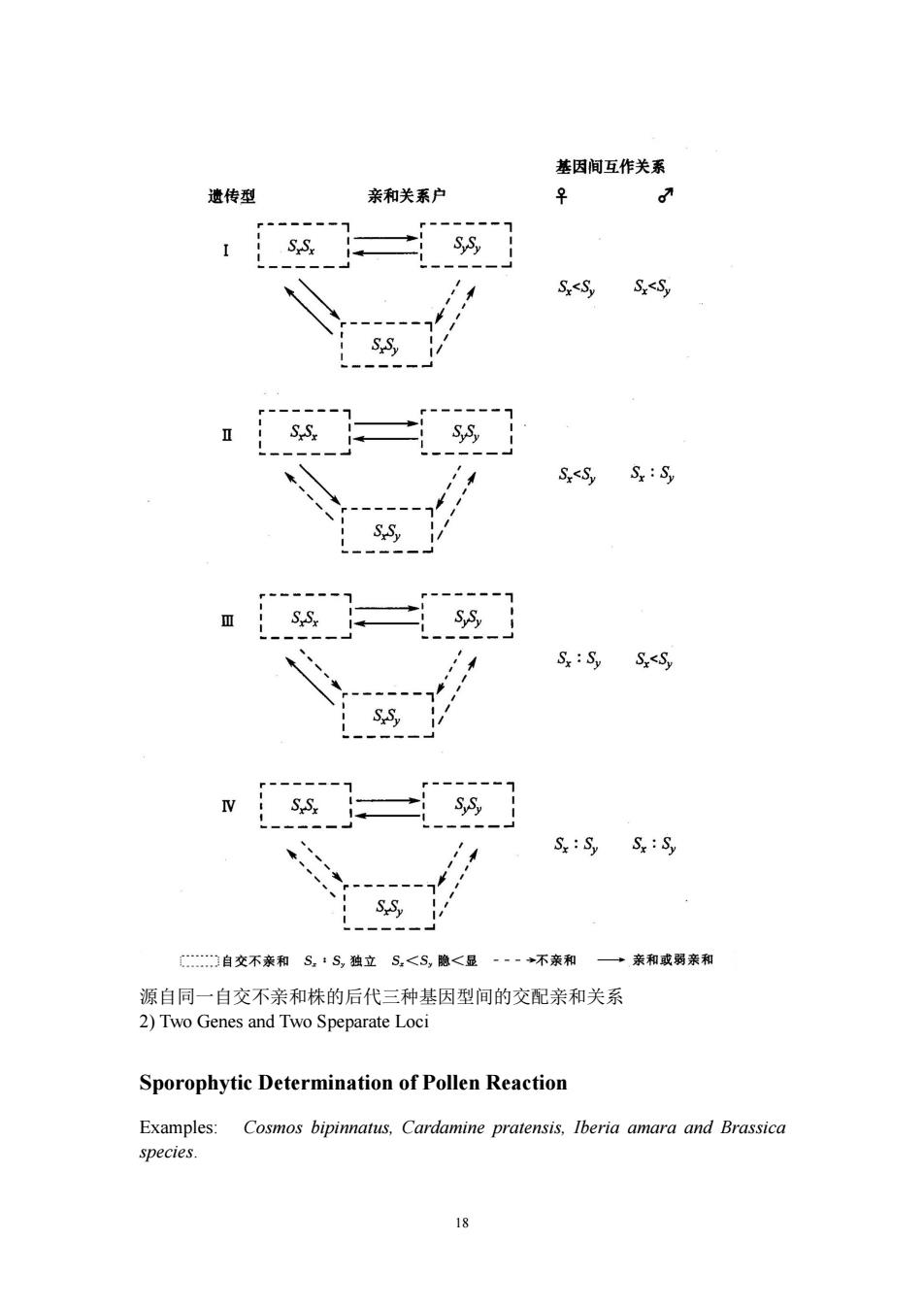
基因间互作关系 遗传型 亲和关系户 ♀ 0 SS, Sx<Sy S.<S S.S Sx<Sy Sy Sy S.S, Sx:Sy S<Sy iS. SS, SySy Sy:Sy S.S, 自交不亲和SS,独立S.<S,隐<显 一一一不亲和 →亲和或弱亲和 源自同一自交不亲和株的后代三种基因型间的交配亲和关系 2)Two Genes and Two Speparate Loci Sporophytic Determination of Pollen Reaction Examples: Cosmos bipinnatus,Cardamine pratensis,Iberia amara and Brassica species. 18
18 源自同一自交不亲和株的后代三种基因型间的交配亲和关系 2) Two Genes and Two Speparate Loci Sporophytic Determination of Pollen Reaction Examples: Cosmos bipinnatus, Cardamine pratensis, Iberia amara and Brassica species

Distinguishing the Sporophtic System from the Gametophytic System 1)Variety of mating patterns 2)Reciprocal differences 3)Incompatibility of female parent 4)Number of incompatibility groups 5)Occurrence of genotypes 6)Number of genotypes 7)Effect of polyploidy upon the action of self-incompatibility mechanism 8)Time of S-allele action 2.Physiology of Self-Incompatility Incompatibility alleles are parent in both the female and male tissure,that is,ovule, ovary,pollen grain and pollen tube.It is important to note that self-incompatibility reaction takes place only when identical genetic information is present in both the male and female tissues,the same allele acting in different ways in pollen and megasporangial tissues.Several theories have been advanced on the physiological and chemical nature of self-incompatibility. East's Immunological Theory East,1929):This theory assumes that incompatibility is due to a positive inhibitory reaction rather than absence of a specific stimulant since pollen grains are capable of germinating and growing in a laboratory medium and also in styles of different species.The inhibitory reaction is highly specific in the sence that only pollen and styles with like alleles react together. The secretions of the pollen tube act as antigens against the stylar tissue which forms antibodies against such a pollen tube and in this way inhibits growth. Straub's Theory Straub,1946,1947):This theory envisages a substance is formed in the pollen grain in a definite amount and with a specific quality determined by the S-allele present.This substance(PGF)is regarded as necessary for the growth of the pollen tube in the style and probably because of its enzyme-like property,is able to decompose the stylar tissue.When all the substance is consumed in growth, the tubes do not grow further. From an experimental viewpoint,there are two types of self-incompatible plants: 1)Plants in which incompatibility reaction takes place during growth of the pollen tubes through the styles,for example in Solanaceae,Unagraceae and Scrophulariaceae. 2)Plants in which incompatibility reaction takes place on the surface of the stigma,for example,Cruciferae,Compositeae. 3.Self-incompatibility in Plant Breeding 19
19 Distinguishing the Sporophtic System from the Gametophytic System 1) Variety of mating patterns 2) Reciprocal differences 3) Incompatibility of female parent 4) Number of incompatibility groups 5) Occurrence of genotypes 6) Number of genotypes 7) Effect of polyploidy upon the action of self-incompatibility mechanism 8) Time of S-allele action 2. Physiology of Self-Incompatility Incompatibility alleles are parent in both the female and male tissure, that is, ovule, ovary, pollen grain and pollen tube. It is important to note that self-incompatibility reaction takes place only when identical genetic information is present in both the male and female tissues, the same allele acting in different ways in pollen and megasporangial tissues. Several theories have been advanced on the physiological and chemical nature of self-incompatibility. ◇East’s Immunological Theory ( East, 1929): This theory assumes that incompatibility is due to a positive inhibitory reaction rather than absence of a specific stimulant since pollen grains are capable of germinating and growing in a laboratory medium and also in styles of different species. The inhibitory reaction is highly specific in the sence that only pollen and styles with like alleles react together. The secretions of the pollen tube act as antigens against the stylar tissue which forms antibodies against such a pollen tube and in this way inhibits growth. ◇Straub’s Theory ( Straub, 1946, 1947): This theory envisages a substance is formed in the pollen grain in a definite amount and with a specific quality determined by the S-allele present. This substance (PGF) is regarded as necessary for the growth of the pollen tube in the style and probably because of its enzyme-like property, is able to decompose the stylar tissue. When all the substance is consumed in growth, the tubes do not grow further. From an experimental viewpoint, there are two types of self-incompatible plants: 1) Plants in which incompatibility reaction takes place during growth of the pollen tubes through the styles, for example in Solanaceae, Unagraceae and Scrophulariaceae. 2) Plants in which incompatibility reaction takes place on the surface of the stigma, for example, Cruciferae, Compositeae. 3. Self-incompatibility in Plant Breeding

Genetically controlled self-incompatibility in flowering plants offers unique economic opportunities as described below.It is a potential tool in breeding FI hybrids, synthetics,triploids,etc Production of hybrids(triploids) Production of F1 hybrids Ornamental plants:The blooming period of ornamentals and the vegetative phase in vegetables say leafy and root vegetables)can be prolonged by preventing seed production. Horticultural crops:In many fruit crops,for example apple,the presence of self-incompatibility necessitates mixed plantings which leads to uneven quality. Development of self-fertile strains from self-incompatible ones by inducing stable type mutants of S-alleles conferring self-fertility would be highly desirable.Selfing in these fruit trees would release potential variability and would permit segregation of rare recessive phenotypes in this highly heterozygous material. Limitations of Self-incompatibility 1)To breed FI hybrids development of homozygous lines is necessary.For this repeated selfing of lines must be possible. 2)Production of hybrid seed invoving the homozygous lines produced in the manner outlined earlier is possible,provided the possibilities of self-and sib-fertilization are completely excluded. 20
20 Genetically controlled self-incompatibility in flowering plants offers unique economic opportunities as described below. It is a potential tool in breeding F1 hybrids, synthetics, triploids, etc. Production of hybrids( triploids) Production of F1 hybrids Ornamental plants: The blooming period of ornamentals and the vegetative phase in vegetables ( say leafy and root vegetables) can be prolonged by preventing seed production. Horticultural crops: In many fruit crops, for example apple, the presence of self-incompatibility necessitates mixed plantings which leads to uneven quality. Development of self-fertile strains from self-incompatible ones by inducing stable type mutants of S-alleles conferring self-fertility would be highly desirable. Selfing in these fruit trees would release potential variability and would permit segregation of rare recessive phenotypes in this highly heterozygous material. Limitations of Self-incompatibility 1) To breed F1 hybrids development of homozygous lines is necessary. For this repeated selfing of lines must be possible. 2) Production of hybrid seed invoving the homozygous lines produced in the manner outlined earlier is possible, provided the possibilities of self- and sib-fertilization are completely excluded