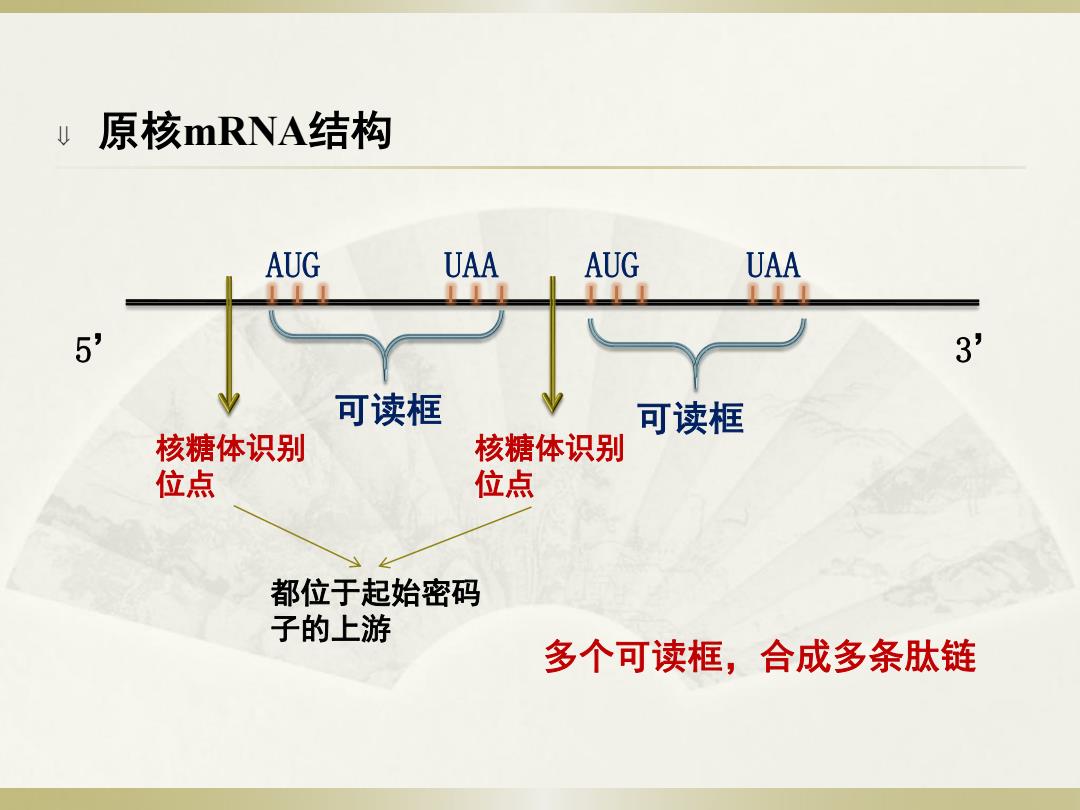
u原核mRNA结构 AUG UAA。 AUG UAA 5' 3 可读框 可读框 核糖体识别 核糖体识别 位点 位点 都位于起始密码 子的上游 多个可读框,合成多条肽链
原核mRNA结构 可读框 AUG UAA 5’ 3’ 可读框 多个可读框,合成多条肽链 核糖体识别 位点 AUG UAA 核糖体识别 位点 都位于起始密码 子的上游
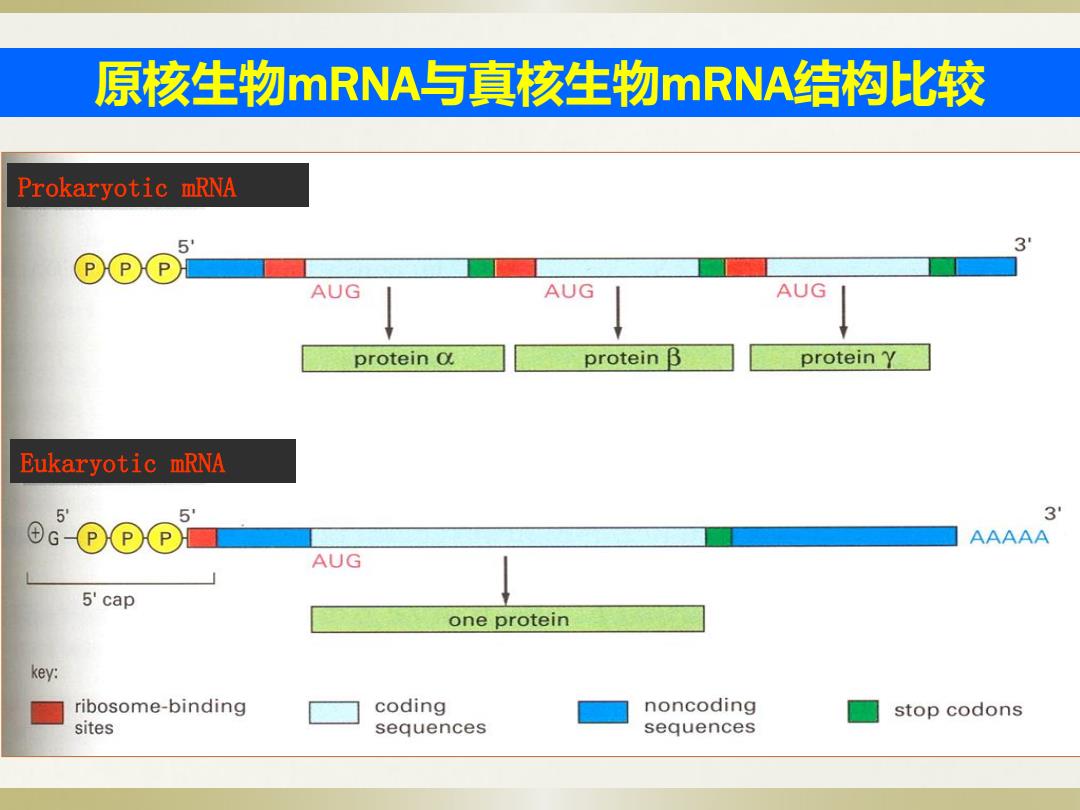
原核生物mRNA与真核生物mRNA结构L比较 Prokaryotic mRNA 5 3 ⊙⊙ AUG AUG AUG protein Q protein B protein Y Eukaryotic mRNA 5 3 AAAAA AUG 5'cap one protein key: ribosome-binding coding noncoding stop codons sites sequences sequences
原核生物mRNA与真核生物mRNA结构比较 核糖体可以不从mRNA上解离连续合成三个蛋白质 Eukaryotic mRNA Prokaryotic mRNA

2、tRNA转运活化的氨基酸至mRNA模板 ,tRNA有两个关键部位:氨基酸结合部位,与 mRNA的结合部位 u 对于组成蛋白质的20种氨基酸,每一种有相应的 tRNA负责转运,aa-tRNA
2、tRNA转运活化的氨基酸至mRNA模板 tRNA有两个关键部位:氨基酸结合部位,与 mRNA的结合部位 对于组成蛋白质的20种氨基酸,每一种有相应的 tRNA负责转运,aa-tRNA

RNA高级结构 1964Holy.R.鉴定出tRNAphe的二级结构为三 叶草形(77个NT) u三叶草结构(5个臂4个环) ua、氨基酸接受臂(aa accept arm) tRNA的5与3?-末端碱基配对形成 一3?端永远为不配对的CCA序列 最后的A的3'或2’OH可以被氨酰化
tRNA高级结构 1964 Holly. R. 鉴定出 tRNAphe 的二级结构为三 叶草形(77个NT) 三叶草结构 (5个臂 4个环) a、氨基酸接受臂(aa accept arm) tRNA 的5’与3’-末端碱基配对形成 3’ 端永远为不配对的CCA序列 最后的A的 3’ 或 2’-OH可以被氨酰化

ub、另外是D(DHU环 双氢脲嘧啶)环D臂 反密码子环反密码子臂(anti一codon arm) 附加臂(extra arm)TyC环TyC臂 u其中 附加臂通常是可变的 uc、含丰富的稀有碱基(约70余种 碱基核 糖残基)反密码子3’端邻近部位..…
b、另外是 D (DHU环 双氢脲嘧啶)环 D臂 反密码子环 反密码子臂(anti—codon arm) 附加臂(extra arm)TψC环 TψC臂 其中 附加臂通常是可变的 c、 含丰富的稀有碱基(约70余种 碱基 核 糖残基)反密码子 3 ’ 端邻近部位………