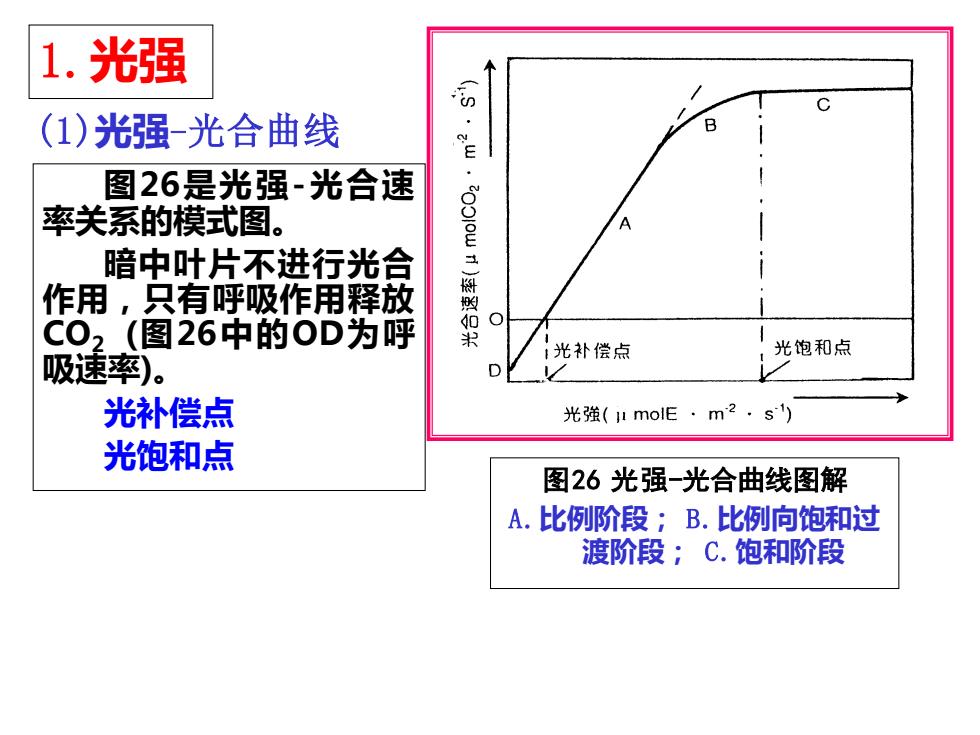
1.光强 图26是光强-光合速 率关系的模式图。 暗中叶片不进行光合 作用,只有呼吸作用释放 CO2 (图26中的OD为呼 吸速率)。 光补偿点 光饱和点 图26 光强-光合曲线图解 A.比例阶段; B.比例向饱和过 渡阶段; C.饱和阶段 (1)光强-光合曲线
1.光强 图26是光强-光合速 率关系的模式图。 暗中叶片不进行光合 作用,只有呼吸作用释放 CO2 (图26中的OD为呼 吸速率)。 光补偿点 光饱和点 图26 光强-光合曲线图解 A.比例阶段; B.比例向饱和过 渡阶段; C.饱和阶段 (1)光强-光合曲线

➢在低光强区,光合速率随 光强的增强而呈比例地增加 (比例阶段,直线A);当超 过一定光强,光合速率增加 就会转慢(曲线B);当达到 某一光强时,光合速率就不 再增加,而呈现光饱和现象。 以后的阶段称饱和阶段(直线 C)。 ➢比例阶段中主要是光强制 约着光合速率,而饱和阶段 中CO2扩散和固定速率是主 要限制因素。 图26 光强-光合曲线图解 A.比例阶段; B.比例向饱 和过渡阶段; C.饱和阶段
➢在低光强区,光合速率随 光强的增强而呈比例地增加 (比例阶段,直线A);当超 过一定光强,光合速率增加 就会转慢(曲线B);当达到 某一光强时,光合速率就不 再增加,而呈现光饱和现象。 以后的阶段称饱和阶段(直线 C)。 ➢比例阶段中主要是光强制 约着光合速率,而饱和阶段 中CO2扩散和固定速率是主 要限制因素。 图26 光强-光合曲线图解 A.比例阶段; B.比例向饱 和过渡阶段; C.饱和阶段
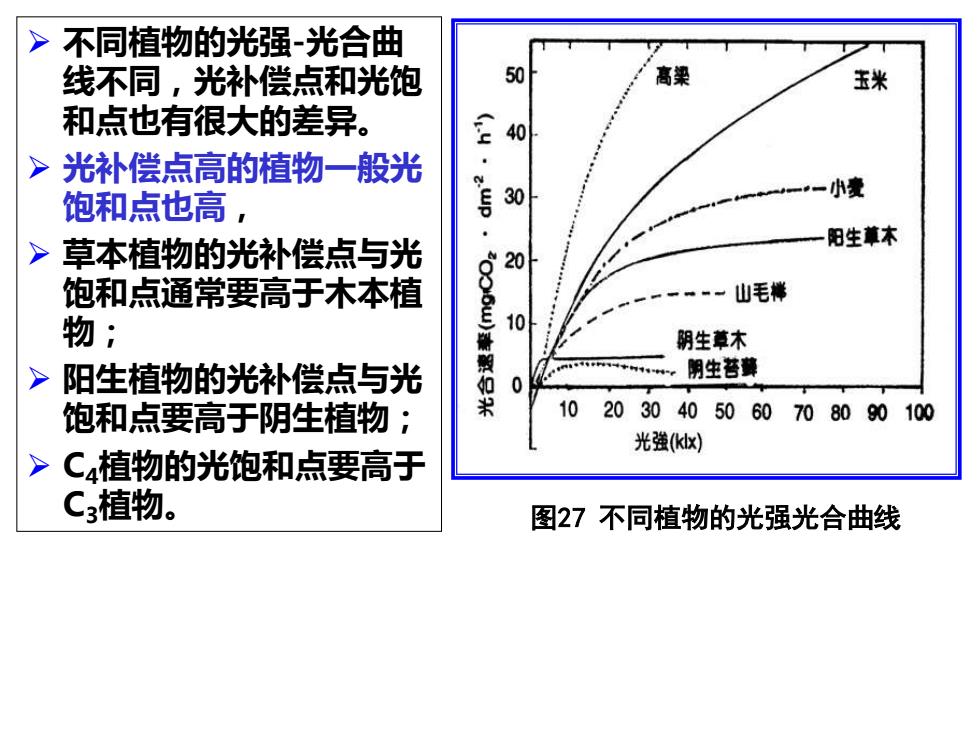
➢ 不同植物的光强-光合曲 线不同,光补偿点和光饱 和点也有很大的差异。 ➢ 光补偿点高的植物一般光 饱和点也高, ➢ 草本植物的光补偿点与光 饱和点通常要高于木本植 物; ➢ 阳生植物的光补偿点与光 饱和点要高于阴生植物; ➢ C4植物的光饱和点要高于 C3植物。 图27 不同植物的光强光合曲线
➢ 不同植物的光强-光合曲 线不同,光补偿点和光饱 和点也有很大的差异。 ➢ 光补偿点高的植物一般光 饱和点也高, ➢ 草本植物的光补偿点与光 饱和点通常要高于木本植 物; ➢ 阳生植物的光补偿点与光 饱和点要高于阴生植物; ➢ C4植物的光饱和点要高于 C3植物。 图27 不同植物的光强光合曲线

(2)强光伤害—光抑制 ➢ 当光合机构接受的光能超过它所能利用的量时,光会引起光 合速率的降低,这个现象就叫光合作用的光抑制。 ➢ 晴天中午的光强常超过植物的光饱和点,很多C3植物,如水 稻、小麦、棉花、大豆、毛竹、茶花等都会出现光抑制,轻 者使植物光合速率暂时降低,重者叶片变黄,光合活性丧失。 ➢ 当强光与高温、低温、干旱等其他环境胁迫同时存在时,光 抑制现象尤为严重。 ➢ 因此光抑制产生的原因及其防御系统引起了人们的重视
(2)强光伤害—光抑制 ➢ 当光合机构接受的光能超过它所能利用的量时,光会引起光 合速率的降低,这个现象就叫光合作用的光抑制。 ➢ 晴天中午的光强常超过植物的光饱和点,很多C3植物,如水 稻、小麦、棉花、大豆、毛竹、茶花等都会出现光抑制,轻 者使植物光合速率暂时降低,重者叶片变黄,光合活性丧失。 ➢ 当强光与高温、低温、干旱等其他环境胁迫同时存在时,光 抑制现象尤为严重。 ➢ 因此光抑制产生的原因及其防御系统引起了人们的重视

光抑制机理 一般认为光抑制主要发生在PSⅡ。按其发生的原初 部位可分为: 受体侧光抑制 常起始于还原型QA的积累。还原型QA的 积累促使三线态P680(P680T)的形成,而P680T可以 与氧作用(P680T +O2→P680 + 1O2 )形成单线态氧 ( 1O2 ); 供体侧光抑制 起始于水氧化受阻。由于放氧复合体不 能很快把电子传递给反应中心,从而延长了氧化型 P680(P680+)的存在时间。 P680+和1O2都是强氧化剂,如不及时消除,它们 都可以氧化破坏附近的叶绿素和D1蛋白,从而使光 合器官损伤,光合活性下降
光抑制机理 一般认为光抑制主要发生在PSⅡ。按其发生的原初 部位可分为: 受体侧光抑制 常起始于还原型QA的积累。还原型QA的 积累促使三线态P680(P680T)的形成,而P680T可以 与氧作用(P680T +O2→P680 + 1O2 )形成单线态氧 ( 1O2 ); 供体侧光抑制 起始于水氧化受阻。由于放氧复合体不 能很快把电子传递给反应中心,从而延长了氧化型 P680(P680+)的存在时间。 P680+和1O2都是强氧化剂,如不及时消除,它们 都可以氧化破坏附近的叶绿素和D1蛋白,从而使光 合器官损伤,光合活性下降